Раздражение, возбудимость и возбуждение у детей
Педагог не только учит и воспитывает детей, но и охраняет их здоровье. Он должен принимать активное участие во всех мероприятиях, которые направлены на оздоровление детских коллективов и создание нормальных условий как окружающей среды, так и всего учебно-воспитательного процесса. Он должен прививать детям гигиенические навыки и повышать санитарною культуру в их домашнем быту.
Пройдет много лет, прежде чем беспомощный младенец станет взрослым человеком. В течение всего этого времени ребенок растет, развивается. Изменяются строение и работа его органов, а также потребности организма, его реакции на условия внешней среды. Для создания наилучших условий роста и развития ребенка, для правильного его воспитания и обучения надо знать особенности его организма; понимать, что полезно для него, что вредно и какие меры следует принимать для укрепления здоровья и поддержания нормального развития. Вот почему в план подготовки работников школьных и дошкольных учреждений включена возрастная анатомия, возрастная физиология и дошкольная гигиена.
Физиология изучает жизненные процессы, протекающие в организме, иными словами, работу, или функции, как отдельных органов, так и всего организма в целом. На основе достижений физиологии были разрешены многие вопросы, связанные с правильной организацией питания и общим оздоровлением условий жизни.
Цель контрольной работы – изучить процессы раздражения, возбудимости и возбуждения у детей.
1 Значение и функциональная деятельность элементов нервной системы
Координация физиологических и биохимических процессов в организме происходит посредством регуляторных систем: нервной и гуморальной. Гуморальная регуляция осуществляется через жидкие среды организма – кровь, лимфу, тканевую жидкость, нервная регуляция – посредством нервных импульсов.
Главное назначение нервной системы заключается в обеспечении функционирования организма как единого целого через взаимосвязь между отдельными органами и их системами. Нервная система осуществляет восприятие и анализ разнообразных сигналов из окружающей среды и от внутренних органов.
Нервный механизм регуляции функций организма более совершенен, нежели гуморальный. Это, во-первых, объясняется быстротой распространения возбуждения по нервной системе (до 100–120 м/с), а во-вторых, тем, что нервные импульсы приходят непосредственно к определенным органам. Однако следует иметь в виду, что вся полнота и тонкость приспособления организма к окружающей среде осуществляются при взаимодействии и нервных, и гуморальных механизмов регуляции.
Общий план строения нервной системы. В нервной системе по функциональному и структурному принципу выделяют периферическую и центральную нервную систему.
Центральная нервная система состоит из головного и спинного мозга. Головной мозг расположен внутри мозгового отдела черепа, а спинной мозг – в позвоночном канале. На разрезе головного и спинного мозга различают участки темного цвета (серое вещество), образованные телами нервных клеток (нейронов), и белого цвета (белое вещество), состоящие из скоплений нервных волокон, покрытых миелиновой оболочкой.
Периферическая часть нервной системы состоит из нервов, например пучков нервных волокон, которые выходят за пределы головного и спинного мозга и направляются к различным органам тела. К ней также относят любые скопления нервных клеток вне спинного и головного мозга, такие как нервные узлы, или ганглии.
Нейрон (от греч. neuron – нерв) – основная структурная и функциональная единица нервной системы. Нейрон – это сложно устроенная высокодифференцированная клетка нервной системы, функцией которой является восприятие раздражения, переработка раздражения и передача его к различным органам тела. Нейрон состоит из тела клетки, одного длинного маловетвящегося отростка – аксона и нескольких коротких ветвящихся отростков – дендритов.
Аксоны бывают различной длины: от нескольких сантиметров до 1–1,5 м. Конец аксона сильно ветвится, образуя контакты со многими клетками.
Дендриты – короткие сильноветвящиеся отростки. От одной клетки может отходить от 1 до 1000 дендритов.
В различных отделах нервной системы тело нейрона может иметь различную величину (диаметром от 4 до 130 мк) и форму (звездчатую, округлую, многоугольную). Тело нейрона покрыто мембраной и содержит, как и все клетки, цитоплазму, ядро с одним или несколькими ядрышками, митохондрии, рибосомы, аппарат Гольджи, эндоплазматическую сеть.
Возбуждение по дендритам передается от рецепторов или других нейронов к телу клетки, а по аксону сигналы поступают к другим нейронам или рабочим органам. Установлено, что от 30 до 50 % нервных волокон передают информацию в центральную нервную систему от рецепторов. На дендритах имеются микроскопических размеров выросты, которые значительно увеличивают поверхность соприкосновения с другими нейронами.
Нервное волокно. За проведение нервных импульсов в организме отвечают нервные волокна. Нервные волокна бывают:
а) миелинизированные (мякотные); чувствительные и двигательные волокна этого типа входят в состав нервов, снабжающих органы чувств и скелетную мускулатуру, а также участвуют в деятельности вегетативной нервной системы;
б) немиелинизированные (безмякотные), принадлежат в основном симпатической нервной системе.
Миелин выполняет изолирующую функцию и имеет слегка желтоватый цвет, поэтому мякотные волокна выглядят светлыми. Миелиновая оболочка в мякотных нервах через промежутки равной длины прерывается, оставляя открытыми участки осевого цилиндра – так называемые перехваты Ранвье.
Безмякотные нервные волокна не имеют миелиновой оболочки, они изолированы друг от друга только шванновскими клетками (миелоцитами).
2 Возрастные изменения морфофункциональной организации нейрона
На ранних стадиях эмбрионального развития нервная клетка имеет большое ядро, окруженное незначительным количеством цитоплазмы. В процессе развития относительный объем ядра уменьшается. Рост аксона начинается на третьем месяце внутриутробного развития. Дендриты вырастают позже аксона. Синапсы на дендритах развиваются после рождения.
Миелинизация нервных волокон. Уже на ранних стадиях внутриутробного развития аксоны нервных клеток окружены клетками-спутниками, которые образуют своеобразную оболочку. Аксон, окруженный такой оболочкой, называется нервным волокном. На 4-5-м месяце в корешках спинномозговых нервов волокна постепенно приобретают отчетливый белый цвет. Объясняется это образованием особого жироподобного вещества - миелина. Он образуется в клетках-спутниках, которые обтекают аксон, многократно обертывая его тонким слоем своего все удлиняющегося тела. Так возникает миелиновая оболочка нервного волокна. Каждые 1-2 мм она прерывается, образуя перехваты. Миелиновую оболочку можно рассматривать как хорошую изоляцию нервного волокна. Кроме того, в миелинизированных волокнах скорость проведения возбуждения в 10-20 раз больше, чем в волокнах, не покрытых миелином. Это объясняется скачкообразным распространением возбуждения: оно перескакивает от одного перехвата к другому.
Миелинизация нервных волокон как в центральной нервной системе, так и в периферической происходит очень интенсивно в последние месяцы внутриутробного развития. У новорожденного миелинизация нервных волокон спинного мозга и ствола головного мозга почти завершена. В значительной мере миелинизированы волокна черепно-мозговых и спинномозговых нервов.
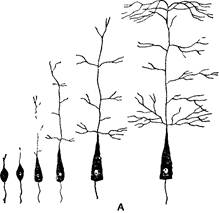
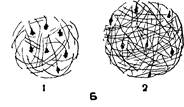
Рис. 1. Развитие нейронов
А — рост пирамидной клетки коры больших полушарий и разрастание дендритов; Б— расстояние между соседними нервными клетками у новорожденного (/), у двухлетнего ребенка (2),
Миелинизация продолжается и после рождения, заканчиваясь в основном к 2-3 годам жизни.
Как правило, миелинизация ускоряется в тех группах волокон, которые начинают усиленно функционировать. Этим объясняется более ранняя миелинизация у недоношенных младенцев. При хронических заболеваниях, связанных с ослаблением двигательной активности, Миелинизация волокон двигательных нервов может значительно задерживаться.
Миелинизация пирамидного пути, проходящего от двигательной области коры больших полушарий до двигательных клеток передних рогов серого вещества спинного мозга, начинается еще до рождения, а с 3-го месяца жизни почти приостанавливается. Лишь примерно с 8-го месяца, в связи с появлением первых попыток ходьбы, интенсивность миелинизации снова, и притом значительно, увеличивается. Миелинизация речевых центров коры в основном заканчивается к 1-2 годам, когда появляется речь.
Очень поздно (не ранее 2-го месяца жизни) начинается миелинизация тех волокон клеток коры больших полушарий, которые идут от одного участка коры к другому. Миелинизируются они очень постепенно, по мере усложнения высшей нервной деятельности. По-видимому, этот процесс прекращается лишь к старости. Особенно медленно указанные волокна получают миелиновую оболочку в лобной области коры, связанной с наиболее сложными проявлениями высшей нервной деятельности.
Функциональные особенности нервных клеток. У новорожденных процессы, протекающие в нервных клетках, замедлены: медленнее возникает возбуждение, медленнее оно распространяется по нервным волокнам. Длительное или сильное раздражение нервной клетки легко приводит ее в состояние торможения. Скорость проведения возбуждения увеличивается по мере миелинизации волокон и к 2-3 годам становится примерно такой же, как и у взрослых. Скорость возникновения возбуждения увеличивается более постепенно и достигает величины, характерной для взрослых лишь к 10—12 годам. Неспособность нервных клеток длительное время находиться в состоянии возбуждения очень характерна и для детей дошкольного возраста. С этим связана нестойкость доминант: любое стороннее раздражение легко разрушает доминанту, вызывая образование нового доминантного очага, который, в свою очередь, быстро оказывается заторможенным. Отсюда неустойчивость внимания дошкольника, быстрый переход от одной деятельности к другой.
Явления иррадиации и индукции. У детей грудного возраста возбуждение легко иррадиирует. Любые рефлекторные движения обычно захватывают значительную часть мускулатуры. Так, движения рук сопровождаются заметной подвижностью ног. Всякое более или менее значительное раздражение вызывает общую двигательную активность. Крику ребенка также сопутствуют движения всего тела. Рефлекторное смыкание век, например при появлении яркого света, сопровождается сжатием губ, а нередко и сгибанием конечностей. При удивлении или при внимательном разглядывании нового предмета ребенок старшего грудного возраста широко открывает не только глаза, но и рот, растопыривая при этом пальцы. Такие иррадиированные реакции характерны и для детей второго года жизни.
В последующие годы устойчивость нервных клеток повышается. Увеличивается сила процессов возбуждения и торможения, в связи с чем более заметными становятся явления индукции: появление очага возбуждения сопровождается понижением возбудимости или торможением других участков мозга. Таким образом, создается препятствие для чрезмерной иррадиации возбуждения. Развитию явлений индукции способствует обучение ходьбе и другим более сложным двигательным актам. При сильном возбуждении, в частности при проявлении радости или огорчения, сохраняется резкая выраженность явлений иррадиации: ребенок прыгает или топает ножками; он весь во власти возбуждения, и никакие уговоры не могут его успокоить.
Дальнейшее усиление явлений взаимной индукции и связанная с этим большая концентрация процессов возбуждения и торможения создают предпосылки для усидчивой целенаправленной деятельности. Этому способствуют воспитание и обучение как в дошкольном, так и в младшем школьном возрасте.
Нервы. Нерв – это скопление нервных волокон, покрытое сверху соединительно-тканной оболочкой. Нерв, передающий возбуждение из центральной нервной системы к иннервируемому органу (эффектору), называют центробежным, или эфферентным. Нерв, передающий возбуждение в направлении центральной нервной системы, называют центростремительным, или афферентным.
Большинство нервов – смешанные, в их состав входят как центростремительные, так и центробежные волокна.
Раздражимость. Раздражимостью называют способность живых систем под влиянием раздражителей переходить из состояния физиологического покоя в состояние активности, т. е. к процессу движения, образования различных химических соединений.
Различают раздражители физические (температура, давление, свет, звук), физико-химические (изменение осмотического давления, активной реакции среды, электролитного состава, коллоидного состояния) и химические (химические вещества пищи, химические соединения, образующиеся в организме, – гормоны, продукты обмена веществ и т. п.).
Естественными раздражителями клеток, вызывающими их деятельность, являются нервные импульсы.
Возбудимость. Клетки нервной ткани, как и клетки мышечной ткани, обладают способностью быстро отвечать на раздражение, поэтому такие клетки получили название возбудимых. Способность клеток отвечать на воздействие внешних и внутренних факторов (раздражителей) называют возбудимостью. Мерой возбудимости является порог раздражения, т. е. та минимальная сила раздражителя, которая вызывает возбуждение.
Возбуждение способно распространяться из одной клетки в другую и перемещаться из одного места клетки в другое.
Возбуждение характеризуется комплексом химических, функциональных, физико-химических, электрических явлений. Обязательным признаком возбуждения является изменение электрического состояния поверхностной клеточной мембраны.
3 Свойства импульсов возбуждения в центральной нервной системе. Биоэлектрические явления
Главной причиной возникновения и распространения возбуждения является изменение электрического заряда на поверхности живой клетки, т. е. так называемые биоэлектрические явления.
По обе стороны поверхностной клеточной мембраны в состоянии покоя создается разность потенциалов, равная около -60-(-90) мВ, причем поверхность клетки заряжена электроположительно по отношению к цитоплазме. Эту разность потенциалов называют потенциалом покоя, или мембранным потенциалом. Величина мембранного потенциала для клеток различных тканей различна: чем выше функциональная специализация клетки, тем она больше. Например, для клеток нервной и мышечной тканей она составляет -80-(-90) мВ, для эпителиальной ткани -18-(-20) мВ.
Причиной возникновения биоэлектрических явлений служит избирательная проницаемость клеточной мембраны. Внутри клетки в цитоплазме ионов калия больше в 30–50 раз, чем вне клетки, ионов натрия меньше в 8-10 раз, в 50 раз меньше ионов хлора. В состоянии покоя клеточная мембрана более проницаема для ионов калия, чем для ионов натрия, и ионы калия выходят через поры в мембране наружу. Миграция положительно заряженных ионов калия из клетки сообщает наружной поверхности мембраны положительный заряд. Таким образом, поверхность клетки в покое несет положительный заряд, тогда как внутренняя сторона мембраны оказывается заряженной отрицательно за счет ионов хлора, аминокислот и других органических ионов, которые через мембрану практически не проникают.
Когда участок нервного или мышечного волокна подвергается действию раздражителя, в этом месте возникает возбуждение, проявляющееся в быстром колебании мембранного потенциала, называемого потенциалом действия.
Потенциал действия возникает из-за изменения ионной проницаемости мембраны. Происходит повышение проницаемости мембраны для катионов натрия. Ионы натрия поступают внутрь клетки под действием электростатических сил осмоса, тогда как в покое клеточная мембрана была малопроницаемой для этих ионов. При этом приток положительно заряженных ионов натрия из внешней среды клетки в цитоплазму значительно превышает поток ионов калия из клетки наружу. В результате происходит изменение мембранного потенциала (понижение мембранной разности потенциалов, а также возникновение разности потенциалов противоположного знака – фаза деполяризации). Внутренняя поверхность мембраны стала заряженной положительно, а наружная вследствие потери положительно заряженных ионов натрия – отрицательно, в этот момент регистрируется пик потенциала действия. Потенциал действия возникает в тот момент, когда деполяризация мембраны достигает критического (порогового) уровня.
Повышение проницаемости мембраны для ионов натрия продолжается короткое время. Затем в клетке возникают восстановительные процессы, приводящие к понижению проницаемости мембраны для ионов натрия и возрастанию для ионов калия. Поскольку ионы калия также заряжены положительно, то их выход из клетки восстанавливает исходные отношения потенциалов снаружи и внутри клетки (фаза реполяризации).
Изменение ионного состава внутри клетки и вне ее достигается несколькими путями: активным и пассивным трансмембранным переносом ионов. Пассивный транспорт обеспечивают присутствующие в мембране поры и селективные (избирательные) каналы для ионов (натрия, калия, хлора, кальция). Эти каналы обладают воротной системой и могут быть закрыты или открыты. Активный транспорт осуществляется по принципу натрийкалиевого насоса, который работает, потребляя энергию АТФ. Его основным компонентом является мембранная NA, КАТФаза.
Проведение возбуждения. Проведение возбуждения обусловлено тем, что потенциал действия, возникший в одной клетке (или в одном из ее участков), становится раздражителем, вызывающим возбуждение соседних участков.
В мякотных нервных волокнах миелиновая оболочка обладает сопротивлением и препятствует току ионов, т. е. она выполняет роль электрического изолятора. В миелинизированных волокнах возбуждение возникает только в участках, не покрытых миелиновой оболочкой, – так называемых перехватах Ранвье. Возбуждение в мякотных волокнах распространяется скачкообразно от одного перехвата Ранвье к другому. Оно как бы «перепрыгивает» через участки волокна, покрытые миелином, в результате чего такой механизм распространения возбуждения получил название сальтаторного (от итал. salto – прыжок). Этим объясняется большая скорость проведения возбуждения по мякотным нервным волокнам (до 120 м/с).
По безмякотным нервным волокнам возбуждение распространяется медленно (от 1 до 30 м/с). Это связано с тем, что биоэлектрические процессы клеточной мембраны проходят на каждом участке волокна, по всей его длине.
Существует определенная зависимость между скоростью проведения возбуждения и диаметром нервного волокна: чем толще волокно, тем больше скорость проведения возбуждения.
Передача возбуждения в синапсах. Синапсом (от греч. synapsis – соединение) называется область контакта двух клеточных мембран, обеспечивающих переход возбуждения с нервных окончаний на возбуждаемые структуры. Возбуждение от одной нервной клетки к другой – процесс однонаправленный: импульс передается всегда с аксона одного нейрона на тело клетки и дендриты другого нейрона.
Аксоны большинства нейронов сильно ветвятся на конце и образуют многочисленные окончания на телах нервных клеток и их дендритах, а также на мышечных волокнах и на клетках желез. Количество синапсов на теле одного нейрона может достигать 100 и больше, а на дендритах одного нейрона – нескольких тысяч. Одно нервное волокно может образовать более 10 тыс. синапсов на многих нервных клетках.
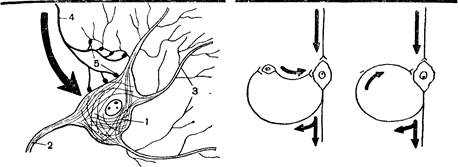
Рис. 2. Схема синапсов
1—тело нервной клетки; 2—ее аксон; 3 — ее дендриты; 4 — аксон другой нервной клетки; 5 — синапсы.
Синапс имеет сложное строение. Он образован двумя мембранами – пресинаптической и постсинаптической, между которыми есть синаптическая щель. Пресинаптическая часть синапса находится на нервном окончании, постсинаптическая мембрана – на теле или дендритах нейрона, к которому передается нервный импульс. В пресинаптической области всегда наблюдаются большие скопления митохондрий.
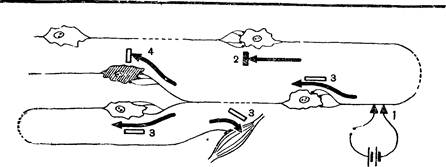
Рис. 3. Схема прохождения импульсов с одного нейрона на другой
Возбуждение через синапсы передается химическим путем с помощью особого вещества – посредника, или медиатора, находящегося в синаптических пузырьках, расположенных в пресинаптической терминали. В разных синапсах вырабатываются разные медиаторы. Чаще всего это ацетилхолин, адреналин или норадреналин.
Выделяют также электрические синапсы. Они отличаются узкой синаптической щелью и наличием поперечных каналов, пересекающих обе мембраны, т. е. между цитоплазмами обоих клеток есть прямая связь. Каналы образованы белковыми молекулами каждой из мембран, соединенных комплементарно. Схема передачи возбуждения в таком синапсе подобна схеме передачи потенциала действия в гомогенном нервном проводнике.
В химических синапсах механизм передачи импульса следующий. Приход нервного импульса в пресинаптическое окончание сопровождается синхронным выбросом в синаптическую щель медиатора из синаптических пузырьков, расположенных в непосредственной близости от нее. Обычно в пресинаптическое окончание приходит серия импульсов, частота их возрастает при увеличении силы раздражителя, приводя к увеличению выделения медиатора в синаптическую щель. Размеры синаптической щели очень малы, и медиатор, быстро достигая постсинаптической мембраны, взаимодействует с ее веществом. В результате этого взаимодействия структура постсинаптической мембраны временно изменяется, проницаемость ее для ионов натрия повышается, что приводит к перемещению ионов и, как следствие, возникновению возбуждающего постсинаптического потенциала. Когда этот потенциал достигает определенной величины, возникает распространяющееся возбуждение – потенциал действия. Через несколько миллисекунд медиатор разрушается специальными ферментами.
Выделяют также особые синапсы тормозного действия. Полагают, что в специализированных тормозящих нейронах, в нервных окончаниях аксонов вырабатывается особый медиатор, оказывающий тормозящее воздействие на последующий нейрон. В коре больших полушарий головного мозга таким медиатором считают гамма-аминомасляную кислоту. Структура и механизм работы синапсов тормозного действия аналогичны таковым у синапсов возбуждающего действия, только результатом их действия является гиперполяризация. Это ведет к возникновению тормозного постсинаптического потенциала, в результате чего наступает торможение.
На каждой нервной клетке расположено множество возбуждающих и тормозящих синапсов, что создает условия для различных ответов на прошедшие сигналы.
4 Процессы возбуждения и торможения в центральной нервной системе
Возбуждение и торможение не самостоятельные процессы, а две стадии единого нервного процесса, они всегда идут друг за другом.
Если возникло возбуждение в определенной группе нейронов, то вначале оно распространяется на соседние нейроны, т. е. происходит иррадиация нервного возбуждения. Затем возбуждение концентрируется в одном пункте. После этого вокруг группы возбужденных нейронов возбудимость падает, и они приходят в состояние торможения, происходит процесс одновременной отрицательной индукции.
В нейронах, которые были возбуждены, после возбуждения обязательно возникает торможение, и наоборот, после торможения в тех же нейронах появляется возбуждение. Это последовательная индукция. Если вокруг групп заторможенных нейронов возбудимость возрастает и они приходят в состояние возбуждения – это одновременная положительная индукция. Следовательно, возбуждение переходит в торможение, и наоборот. Это значит, что обе эти стадии нервного процесса сопутствуют друг другу.
Заключение
Завершая работу, можно прийти к выводу, что состояние возбуждения, а следовательно, и сокращения мышцы возникает под влиянием волн возбуждения, или импульсов, которые с определенной частотой бегут по нервным волокнам. Каждый импульс вызывает в мышце очередную волну возбуждения. Новая волна возбуждения может возникнуть только по окончании предыдущей. Установлено, что возбудимость мышечных волокон, определяемая той минимальной силой раздражения, которая еще может вызвать в них волну возбуждения, достигает уровня, соответствующего взрослым, уже в первые годы жизни. Но скорость протекания в мышце отдельных волн возбуждения и их частота продолжают увеличиваться до 10 лет. Увеличение скорости протекания волн возбуждения способствует большей мобилизации мышечной силы.
Список литературы
1. Антонова О.А. Возрастная анатомия и физиология. – М., 2008.
2. Гальперин С.И. Анатомия и физиология человека. - М.: Высшая школа, 1974.
3. Малая медицинская энциклопедия: В 6 т. Т. 6. - М.: Медицина, 1991–1996.
4. Матюшонок М.Т., Турин Г.Г., Крюкова А.А. Физиология и гигиена детей и подростков. - М.: Высшая школа, 1974.
5. Хрипкова А.А. Возрастная физиология. - М.: Просвещение, 1978.