Динамика видового разнообразия сфагновых мхов на болотах Западной Сибири по трансекту лесостепь - Тундра
В экосиситемах болот сообщества мхов играют значительную роль, которая проявляется в специфическом сложении растительного покрова, синтезе органического вещества и его депонировании в форме торфяных залежей, а также в развитии самих гидроморфных комплексов.
Одним из важнейших показателей условий местопроизрастания является величина годичного прироста фитомассы как среднего для всего сообщества в целом, так и для видов растений, образующих эколого-ценотические группировки.
Значимость сообществ мхов в структурной организации растительного покрова болот очевидна. Особенно это касается лесоболотных регионов Сибири, где гидроморфные и полугидроморфные комплексы по существу на равных конкурируют с экосистемами суходольного ряда развития, а в отдельных районах и преобладают над ними.
Детальное изучение моховых сообществ в конкретных болотных экосистемах с дифференцированным подходом к выявлению особенностей условий произрастания и строения моховых микроценозов является базовой основой для экстраполяции скорости накопления и количества аккумулируемого углерода на большие заболоченные территории (1).
Все вышесказанное обусловливает актуальность проблемы, изучению которой посвящена настоящая дипломная работа.
Под природопользованием понимается целенаправленная совокупность видов деятельности, задача которых сводится к обеспечению потребностей настоящего и будущего поколения людей в качестве и разнообразии природной среды, улучшению использования и воспроизводства природных ресурсов, сохранению равновесия между природой и обществом на основе взаимосвязанных мер по охране, воспроизводству и рациональному потреблению природного потенциала.
Болото - избыточно увлажненный участок земной поверхности, для которого характерно постоянное переувлажнение и дефицит кислорода, произрастание особой влагостойкой растительности и накопление частично разложившегося органического вещества, превращающегося в дальнейшем в торф, слоем не менее 30 см. При глубине торфа менее 30 см участок относится к заболоченным землям. Торф накапливается в результате болотообразовательного процесса. Иногда считают, что термин "заболачивание" является синонимом процесса болотообразования. Последний, однако, шире и включает в себя первый. Заболачивание - это только начальная стадия возможного образования болота и для нее характерна двойственность проявления, заключающаяся в обратимости процессов заболачивания - разболачивания. Ежегодно в мире заболачивается около- 660 км2 земли. В процессе развития болотообразования формируется торфяная залежь, достигающая разной мощности с наибольшими значениями - 12-15 м. 'Болото - экосистема, состоящая из трех основных компонентов: воды, специфической бойотной растительности и торфа, и поэтому болото является предметом внимания нескольких самостоятельных направлений (2). Ботаники и геоботаники изучают в них индивидуальность болотной растительности, а по стратиграфии торфяных залежей - климатические характеристики периода торфонакопления и определяют их как болота. Геологи определяют запасы в границах промышленных залежей и называют торфяные болота торфяными месторождениями. Лесники изучают болота с позиций улучшения бонитета древостоя и называют их лесными болотами, а почвоведы — с позиций получения сельскохозяйственных угодий и называют их торфяными почвами на органогенных породах. Разночтения в понятиях торфяные месторождения, торфяные болота, заболоченные земли, торфяные почвы проявляются в дальнейшем расхождениями в количественных оценках площадей болот.
1. ЗАПАДНАЯ СИБИРЬ. КРАТКАЯ ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА
Границы и площадь. Западная Сибирь представляет собой территорию, простирающуюся на 2500 км от Северного Ледовитого океана до сухих степей Казахстана и на 1500 км от гор Урала до Енисея. Около 80% площади Западной Сибири расположено в пределах Западно-Сибирской равнины, которая состоит из двух плоских чашеобразных сильно заболоченных впадин, разделенных повышенными до 175-200 м Сибирскими Увалами. Общая площадь Западной Сибири составляет 2.4 млн. км2 (3).
Геология и орография. В основании Западно-Сибирской равнины лежит Западно-сибирская плита. В основании Западно-сибирской плиты находится палеозойский фундамент, глубина залегания которого составляет, в среднем, 7 км.
Образование Западно-сибирской плиты началось в верхней юре, когда в результате обламывания, разрушения и перерождения огромная территория между Уралом и Сибирской платформой опустилась, и возник огромный седиментационный бассейн. В ходе своего развития Западно-сибирская плита не раз захватывалась морскими трансгрессиями. В конце нижнего олигоцена море покинуло Западно-сибирскую плиту, и она превратилась в огромную озерно-аллювиальную равнину. В среднем и позднем олигоцене и неогене северная часть плиты испытала поднятие, которое в четвертичное время сменилось опусканием. Общий ход развития плиты с опусканием колоссальных пространств напоминает не дошедший до конца процесс океанизации. Эта особенность плиты подчеркивается феноменальным развитием заболоченности.
Климат. Западная Сибирь находится почти на одинаковом расстоянии как от Атлантического океана, так и от центра континентальности Евразии, поэтому ее климат носит умеренно континентальный характер. Зимой и в летнее время, когда циклоническая деятельность, а с ней и поступление атлантического воздуха ослабевают, в Западную Сибирь поступает арктический воздух. Глубокому проникновению арктических воздушных масс способствует равнинность местности и открытость ее к северу.
Средняя температура января уменьшается от -15ОС на юго-западе до -30 ОС на северо-востоке Западной Сибири. Средняя температура июля увеличивается от +5 ОС на севере до +20 ОС на юге. Наибольшей континентальностью отличается северо-восток Западной Сибири, где разности средних температур января и июля достигают 45 О.
Гидрография. Реки Западной Сибири принадлежат бассейну Карского моря. Самая крупная водная артерия - Обь с притоком Иртыш - относится к числу величайших рек земного шара. Река Обь образуется при слиянии Бии и Катуни, берущих начало на Алтае, и впадает в Обскую губу Карского моря. Среди рек России она занимает первое место по площади бассейна и третье по водности. В лесной зоне, до устья Иртыша, Обь принимает свои основные притоки: справа - реки Томь, Чулым, Кеть, Тым, Вах; слева - реки Парабель, Васюган, Большой Юган и Иртыш. Наиболее крупные реки севера Западной Сибири - Надым, Пур и Таз - берут свое начало на Сибирских Увалах.
Географическое зонирование. Западно-Сибирская равнина в природном отношении характеризуется достаточно ярко выраженной широтной зональностью. На крайнем севере полуостровной части равнины раскинулись тундры, материковую часть занимает лесотундра. Южные районы относятся к лесостепной и степной природным зонам. В северных районах равнины очень широким распространением пользуются болота. На их долю приходится более половины общей площади (4).
Тундра, занимающая самую северную часть Тюменской области (п-ова Ямал и Гыданский) и имеющая площадь около 160 тыс. км2, не имеет лесов. Лишайниковые и моховые тундры Западной Сибири встречаются в сочетании с гипново-травяными и лишайниково-сфагновыми, а также крупнобугристыми болотными массивами.
Зона лесотундры простирается к югу от тундры полосой примерно 100-150 км. Как переходная зона между тундрой и тайгой она представляет собой мозаичное сочетание участков редколесий, болот, зарослей кустарников. Лесная (таежная, лесоболотная) зона охватывает пространство между 66О и 56Ос.ш. полосой примерно в 1000 км. В нее входят северная и средняя части Тюменской области, Томская область, северная часть Омской и Новосибирской областей, занимая около 62% территории Западной Сибири.
Лесную зону Западно-Сибирской равнины подразделяют на подзоны северной, средней, южной тайги и березово-осиновых лесов. Основным типом лесов зоны являются темнохвойные леса с преобладанием ели сибирской, пихты сибирской и сосны сибирской (кедра). Темнохвойные леса встречаются почти всегда лентами по долинам рек, где они находят условия необходимого для них дренажа. На водоразделах они приурочены только к холмистым, возвышенным местам, а плоские территории заняты преимущественно болотами (5).
Важнейший элемент ландшафтов тайги - болота низинного, переходного и верхового типа. Лесистость Западной Сибири составляет всего 30.5% и является следствием слабой расчлененности и связанной с ней слабой дренированности всей территории региона, что способствует развитию не лесообразовательных, а болотообразовательных процессов на всей площади таежной зоны. Западно-Сибирская равнина характеризуется исключительной обводненностью и заболоченностью, ее средняя и северная части относятся к одним из самых переувлажненных пространств на земной поверхности. Самые крупные в мире болотные массивы (Васюганский) расположены в южной тайге (6). Наряду с темнохвойной тайгой на Западно-Сибирской равнине встречаются сосновые леса, приуроченные к песчаным наносам древних аллювиальных равнин и к песчаным террасам вдоль речных долин. Кроме того, в пределах лесной зоны сосна является характерным деревом сфагновых болот и образует своеобразные ассоциации сфагновых сосняков на заболоченных почвах.
Лесостепная зона, примыкающая к подзоне лиственных лесов лесной зоны, характеризуется присутствием и лесных, и степных растительных сообществ, а также болот (рямов), солончаков и лугов. Характерной чертой лесостепи Западной Сибири является гривно-лощинный рельеф и обилие соленых бессточных озер.
Болота Западной Сибири. Общая заболоченность территории Западно-Сибирской равнины площадью почти 3 млн. км2 в среднем составляет 50%, достигая в отдельных речных бассейнах 70-80%. Процесс образования болот начался 10-12 тыс. лет назад в условиях резко континентального климата позднеледниковья. В дальнейшем, в условиях постоянного избыточного увлажнения и теплого климата атлантического периода (4,5-8,0 тыс. лет тому назад), заболачивание усилилось и превратилось из локального в регионально-локальное. Центры заболачивания, первоначально изолированные, постепенно сливались между собой и превращались в обширные болотные торфяные системы (7). Крупнейшие болота – это Лайменское, занимающее 50,2 тыс. га; Салымо-Юганское -73,9 тыс. га, Озерное Большое – 572,4 тыс. га, Пассал-Когот – 210,3 тыс. га, Большое Васюганское, площадь которого определяется в 5 млн.га. Всего на Западно-Сибирской равнине образовалось 5 тыс. торфяных месторождений общей площадью более 30 млн. га и запасами торфа 113,7 млрд. т (40% от запасов в мире).
2. МОХООБРАЗНЫЕ БОЛОТНЫХ ЭКОСИСТЕМ
Моховидные (Bryophyta) часто, даже в кругах специалистов, называют популярным и коротким именем – мхи. Однако в более точном, научном смысле собственно мхами называют представителей лишь одной, наиболее обширной группы отдела моховидных, а именно – листостебельные, или настоящие мхи (Bryopsida) (8).
Среди высших растений моховидные образуют наиболее обособленную группу.
Моховидные иногда рассматривают в целом как растения, связанные в своей жизни с достаточным, часто избыточным увлажнением. Значение этой зависимости не следует преувеличивать. Прежде всего, все развитие гаметофита совершается у них в воздушной среде. Моховидные обнаруживают значительную устойчивость к длительному пересыханию и способны произрастать в местах неравномерного и даже весьма кратковременного сезонного увлажнения.
2.1 Мохообразные как особая линия эволюции высших растений
Моховидные представляют собой слепую ветвь развития растений. С давних пор они прочно заняли свое особое место в экономике природы и сохранили его в сложных условиях формирования континентов, изменения климата и растительного покрова. Массовое участие моховидных в растительном покрове земли оказывает существенное воздействие на среду обитания других растений и животных (9).
Общеизвестно ландшафтное значение моховидных в тундровой зоне, особенно в моховых тундрах.
Моховидные в подавляющем большинстве многолетние и только сравнительно редко однолетние растения. Но независимо от продолжительности жизни все они низкорослы.
Моховидные отличаются сравнительно простой организацией.
При наличии сложно устроенных органов размножения у моховидных, по существу, отсутствуют специализированные вегетативные органы.
2.2 Цикл развития моховидных
Особенность чередования поколений у моховидных заключается в том, что гаметофит у них значительно расширяет по сравнению с остальными высшими растениями, сферу своей физиологической деятельности. Наряду с обеспечением полового размножения гаметофит моховидных принимает на себя выполнение основных вегетативных функций – фотосинтеза, водоснабжение, минерального питания целого растения. Спорофит же фактически ограничивается спорообразованием.
Из одноклеточной споры у моховидных вырастает гаметофит. Прежде всего развивается многоклеточное ветвистое нитчатое или пластинчатое образование – протонема, на которой у мхов закладывается несколько или даже много почек.
Из почек в одних группах моховидных вырастают пластинчатые, в других – облиственные «побеги», которые в бриологии называют гаметофорами, поскольку они образуют половые органы. Таким образом, протонема, почки на ней и вырастающие из них гаметофоры представляют собой гаметофит (гаметофазу) моховидных.
На гаметофорах образуются половые органы – женские - архегонии и мужские - антеридии (10).
Оплодотворение яйцеклетки подвижными сперматозоидами происходит при наличии капельно-жидкой воды. Слияние этих гамет и дальнейшее развитие зиготы происходит внутри архегония.
Из зиготы за время от нескольких месяцев до двух лет развивается спорофит, у моховидных обычно называемый спорогоном.
2.3 Класс - листостебельные мхи
Из всех моховидных листостебельные мхи включают наибольшее число видов. Среди них многие отличаются широким распространением, высокой жизненной устойчивостью и большой фитоценотической ролью в растительном покрове Земли. Экологическое своеобразие и особое значение мхов в природных процессахво многом зависит от присущих им групповых форм роста, благодаря чему мхи могут на больших площадях создавать рыхлые или плотные покровы различной мощности.Тем самым мхи активно участвуют в формировании на поверхности континентов многочисленных мощных влагоприемников в виде болот и замоховелых лесов, оказывающих существенное влияние на общую обеспеченность суши влагой (11).
Листостебельные мхи - самый крупный класс моховидных. Он состоит из 700 родов и включает 14500 или больше видов. Представители этого класса на суше встречаются почти повсеместно. Этот класс обычно делят на три подкласса: сфагновые мхи, андреевые мхи и бриевые мхи. Рассмотрим один из основных подклассов этого вида – сфагновые мхи.
2.4 Сфагновые мхи
Название происходит от латинизированного sphagnos — род мха.
Примерно 350 видов мхов, относящихся к единственному роду сфагнум Sphagnum (рисунок 1), составляют четко очерченную группу, очень давно отделившуюся от основной линии эволюции этого отдела (12). Стебли листостебельного гаметофита у них несут мутовки ветвей, часто по пять в узле, более тесно скученные у верхушки растения, так что возникает своего рода головка.

Рисунок 1 - Сфагновый мох
Многолетние, беловато-зеленые, желтоватые, буроватые или красноватые болотные мхи. Стебли мха высотой 5—30 см, ветвистые, ризоидов нет, листья однослойные без жилки, образованы двумя типами клеток: водоносными — крупными, мертвыми, бесцветными и пустыми с волокнами и порами в наружных стенках; и хлорофиллоносными, вытянутыми в длину, узкими, окрашенными, расположенными между водоносными.
Стеблевые и веточные листья различны по форме (языковидные, широкояйцевидные или овальные, равносторонние, заостренные, яйцевидно ланцетные), длиной 0,5—2 мм. Коробочки со спорами шаровидные, с крышечкой. Споры желтые или желтовато-бурые. Спороносят в июле — августе. Растения однодомные и двудомные. Имеют два поколения. Домиирует гаметофит (половое поколение) — облиственное растение с половыми органами — антеридиями, в которых развиваются мужские гаметы — сперматозоиды, и архегониями, в которых развиваются женские гаметы — яйцеклетки.
Бесполое поколение представлено безлистным стебельком; на его верхушке имеются коробочки со спорами. Проросшая спора образует пластинчатый заросток — протонему; из нее развивается растение сфагнума. Сфагнум растет медленно (за год вырастает до 3 см), а снизу стебель отмирает.
Сфагновые мхи – наименее требовательные к минеральному питанию болотные растения. Они довольствуются минеральными веществами, поступающими на поверхность почвы из атмосферы. Эти мхи отличаются очень высокой влагоемкостью, позволяющей некоторым видам сфагнума поглощать воды в 20-30 раз больше своего веса в сухом состоянии.
Высокая влагоемкость сфагнового мха объясняется тем, что ткани его листьев состоят в основном из мертвых гиалиновых клеток, между которыми зажаты очень мелкие живые хлорофиллоносные клетки. Из мертвых гиалиновых клеток состоит и наружный покров (гиалодермис) стеблей и ветвей (рисунок 2).

Рисунок 2 - Строение сфагнового мха
1- общий облик Sphagnum magellanicum; 2- наружные клетки гиалодермиса стебля; 3 – наружные клетки гиалодермиса отстоящей ветки; 4 – часть поперечного среза через стебель; 5- веточный лист; 6- поперечный срез веточного листа; 7- часть ткани веточного листа (темные –хлорофилловые клетки, бесцветные – водоносные клетки с порами (а) и волокнами (б)
Так как сфагновый мох не имеет корней, минеральное питание в виде бедных солями водных растворов воспринимается всей поверхностью растения через гиалиновые клетки. Поселяясь на заболачивающихся местах, сфагнум уже сам, в условиях достаточно влажного климата, усиливает болотообразовательный процесс (13).
Торфяные мхи произрастают на торфяных болотах (низинах, переходных и особенно верховых). Они играют большую роль в зарастании водоемов и в заболачивании лесов и лугов. Побеги этих мхов, нарастая верхушкой и отмирая снизу, постепенно превращаются в торф. Растет по всей территории России.
Свойства. Мхи – растения, не имеющие циркуляторной системы. Они получают влагу из осадков или атмосферы, используя осмотическое давление. Это означает также, что они одновременно поглощают все содержащиеся в окружающей среде вещества, в том числе вредные, не обладая механизмами освобождения от них. Поэтому мхи являются прекрасными индикаторами состояния окружающей среды. Сфагновые мхи способны повышать кислотность окружающей их среды, выделяя в воду ионы водорода. Наиболее важной особенностью сфагнума, приобретенной в ходе миллионов лет эволюции, является его способность впитывать и сохранять от 12 до 20 весовых частей воды на часть сухого веса (в зависимости от биологического вида сфагнума), а также его бактерицидные свойства.
Размножение. Сфагнум может размножаться как спорами, так и вегетативно. Количество спор в спорофите может быть от 20 000 до 200 000 в зависимости от вида мха, а на квадратном метре болота – примерно 15 млн. Спорофит выбрасывает споры в июле. Коробочка как бы взрывается при сухой теплой погоде, и споры разносятся ветром на различные расстояния, так как имеют разный размер, 20-50 мкм. Еще один механизм переноса спор – потоком воды или брызгами от капель дождя. В последнем случае расстояние переноса не превышает десятка сантиметров. Размножение спорами является основным при расселении сфагнума на большие расстояния - новые или пострадавшие от пожара или хозяйственной деятельности участки. Для образования растения из споры необходимо, чтобы она попала на подходящую почву – влажный торф. Другой механизм распространения сфагнума – вегетативный, участками стебля или ветвей. Этот механизм эффективен на малых расстояниях (14).
Места обитания. Основной средой обитания сфагнума в России являются болота, занимающие примерно пятую часть ее территории.
Поверхность моховой дернины очень живописна: на ней видны лишь головки сфагнума всевозможных оттенков, напоминающие узоры персидского ковра (рисунок 3).

Рисунок 3 - Головки сфагновых мхов
У сфагнума одновременно протекают процессы роста и разложения. Верхушка растет, вытягиваясь вверх на 1-3 см в год, а нижняя подводная часть отмирает и со временем превращается в торф, поэтому стебель постепенно опускается вниз. Однако, из-за постоянного накопления торфа (до 1 см в год в верхних слоях) поверхность болота медленно поднимается – формируются так называемые верховые болота, в которых обычно нет трясин, а уровень воды находится на 10-20 см ниже поверхности сфагновой дернины (15).
Отдельные растения мха вместе образуют могучую дернину (рисунок 4).

Рисунок 4 – Дернина сфагновых мхов
Отмирающие нижние части сфагновых мхов формируют многометровые отложения торфа. В верхних слоях идет постепенное разложение органических веществ, нижние под давлением верхних слоев уплотняются на глубине несколько метров, одному году соответствует уже слой толщиной несколько миллиметров, а возраст глубинных слоев составляет несколько тысячелетий.
Анатомия. Гаметофор сфагнов состоит из двух типов клеток – живых, хлорофиллоносных, и мертвых, лишенных содержимого крупных водоносных, или гиалиновых, клеток.
Стенки последних укреплены спиральными волокнами, предохраняющими их от разрыва. У многих видов в стенках гиалиновых клеток имеются поры, облегчающие всасывание воды (рисунок 5).
Листья, произрастающие на веточке, налегают друг на друга, называют – черепитчато-налегающими (16).

Рисунок 5 - Клетки сфагна: 1 – водоносная клетка; 2 – хлорофиллоносная клетка; 3 – пора; 4 – спиральные волокна
Более сложное строение обнаруживает стебель. В нем клетки, хотя и дифференцированы, но не образуют специализированных тканей, как у сосудистых растений.
Спорогон сфагнов, как и у других мхов, состоит из ножки и коробочки. Ножка спорогона очень короткая а коробочка не имеет перистома.
Листостебельное растение (гаметофор) состоит из стебля с веточками или без них. Как на стебле, так и на веточках у бриофитов имеются листья, которые называются соответственно стеблевыми и веточными.
Экологические особенности и распространение. Мхи способны заселять наиболее бедные и влажные местообитания. В умеренной зоне они образуют обширные торфяные болота. Торф – это неразложившиеся спрессованные остатки сфагновых мхов, которые сохраняются практически в неизменном виде столетиями и тысячелетиями. Многие болота таежной зоны имеют возраст от 8 000 до 10 000 лет. Вместе со сфагнами гниению не подвергаются и остатки других болотных растений. Происходит это потому, что торфяные мхи выделяют в окружающую среду целый букет органических кислот (яблочную, лимонную, уксусную, муравьиную и др.).
Но сфагны способны расти не только в местообитаниях с бедным минеральным питанием. Часто их можно встретить и в поймах лесных речек, и на озерах, и у родников (выходах ключевых вод). В таких местообитаниях они не образуют сплошных ковров, а также торфа, так как биохимический состав растений тех видов мхов, которые здесь произрастают, не столь химически агрессивен (17).
2.5 Разновидности сфагновых мхов
Sphagnum angustifolium ( Russ. ex Russ. ) C. Jens – типично болотный вид, облигатно связанный с торфяными болотами на всем протяжении его ареала. Является доминантом мохового покрова в рослых рямах и мезоолиготрофных осоково – сфагновых топей верховых и переходных болот. С высоким постоянством растет на грядах ГМК и в рямах олиготрофных сфагновых болот.
Sphagnum balticum ( Russ. ) Russ.ex C.Jens –типично болотный вид, облигатно связанный с торфяными болотами на всем протяжении его ареала. Растет в сфагновых топях и мочажинах верховых водораздельных болот атмосферного питания, где является одним из основных доминантов и торфообразователей. Встречается часто, по всей территории.
Spagnum compactum DC. in Lam. et DC – характерный болотный вид, облигатно связанный на юго – востоке Западной Сибири с торфяными болотами. Растет в обширных сфагновых топях верховых водораздельных болот атмосферного питания. Предпочитает местообитания с угнетенным сфагновым покровом и доминированием печеночных мхов в местах выхода болотных газов.
Sphagnum fallax ( Klinggr. ) Klinggr – характерный болотный вид, облигатно связанный на юго – востоке Западной Сибири с торфяными болотами. Растет в мезоолиготрофных осоково – сфагновых топях верховых и переходных болот, где является основным доминантом и торфообразователем. Реже поселяется в мезотрофных осоково – сфагновых топях. Встречается часто, по всей территории.
Sphagnum lindbergii Schimp. ex Lindb ( рисунок6) – гипоарктический характерный болотный вид, облигатно связанный на юго – востоке Западной Сибири с торфяными болотами. Растет в обширных топях и мочажинах верховых водораздельных сфагновых болот.

Рисунок 6 - Сфагновый мох Sphagnum lindbergii
Sphagnum fuscum ( Schimp. ) Klinggr (рисунок 7) – характерный болотный вид, облигатно связанный в лесной зоне Западной Сибири с торфяными болотами. Растет на грядах и пушицево – сфагновых топях олиготрофных болотных комплексов и в рямах, являясь основным доминантом и торфообразователем верховых сфагновых болот.

Рисунок 7 - Сфагновый мох Sphagnum fuscum
Sphagnum jensenii Lindb – типично болотный вид, облигатно связанный с торфяными болотами на всем протяжении его ареала. Растет в застойных топях и мочажинах верховых сфагновых болот, где нередко является доминантом мохового яруса. Поселяется также в проточных мезо – олиготрофных и мезотрофных осоково – сфагновых топях верховых и переходных болот. Встречается сравнительно часто, по всей территории.
Sphagnum magellanicum Brid ( рисунок 8) – типично болотный вид, облигатно связанный с торфяными болотами на всем протяжении его ареала. Растет в рослых и типичных рямах, в олиготрофных болотных комплексах и проточных осоково – сфагновых топях верховых сфагновых болот, реже на кочках и грядах в мезотрофных осоково – сфагновых топях и мелколесьях, а также на мезотрофных лесных сфагновых болотах. Встречается часто, по всей территории (18).

Рисунок 8 - Сфагновый мох Sphagnum magellanicum
3. МЕТОДИКА ОТБОРА ОБРАЗЦОВ И МЕТОДЫ ОЦЕНКИ НАДЗЕМНОЙ, ПОДЗЕМНОЙ И ОБЩЕЙ ПРОДУКЦИИ
Общая первичная продукция в болотах слагается из продукции разных фракций: надземной массы трав, кустарничков и кустарников, сфагновых мхов и лишайников, узлов кущения, корневищ и корней трав и кустарничков.
Для определения динамики запасов и продукции этих фракций на ключевом участке закладывалась пробная площадка (100 м2), на которой отбирались пробы растительного вещества. Фитомасса трав и кустарничков срезалась над поверхностью мхов на квадратах размером 0,25 м2. Масса узлов кущения, корневищ и корней трав и кустарничков, расположенных ниже линии среза, считается подземной. Отдельно учитывалась масса мхов. На квадратах, где была срезана надземная фитомасса, отбирались монолиты величиной 1 дм2 по слоям 10 см от поверхности головок мхов до глубины 30 см. Отобранные пробы разбирались на фракции (19).
Фитомассу трав, кустарничков и кустарников разбирали по видам и каждый вид на следующие фракции: зелёные части трав; живые листья текущего года и прошлых лет у кустарничков; побеги кустарничков текущего года; многолетние одревесневшие части кустарничков, которые располагаются выше линии среза. В монолитах отделялись фотосинтезирующие части мхов, их мёртвые части, очёс и торф.
Надземная продукция. Надземная продукция трав принималась равной максимальному за сезон запасу их зеленой фитомассы (ANPt). У листопадных кустарничков продукция оценивалась как сумма максимальной за сезон массы зеленых листьев и массы побегов текущего года (ANPk1). У вечнозеленых кустарников – по максимальному запасу побегов текущего года с расположенными на них листьями (ANPk2).
Продукция сфагновых мхов определялась как произведение плотности (количество стеблей мха в 1 дм2) на массу годового линейного прироста. Для определения линейного прироста сфагнового мха использовался метод «индивидуальных меток», который заключается в том, что стебель растения окольцовывается под головкой мха крючочком из тонкой проволоки (20).
На другой конец проволоки, расположенный на расстоянии 5-10 см от растения, прикрепляется бирка из алюминиевой фольги с номером. Колечко из проволоки свободно лежит на веточках, не травмируя стебель. В каждой ассоциации окольцовывается 70-100 растений. Отбор окольцованных растений проводится через год. У каждого экземпляра измеряется расстояние от отметки под головкой растения до кольца, которое фиксирует положение головки мха в момент постановки опыта
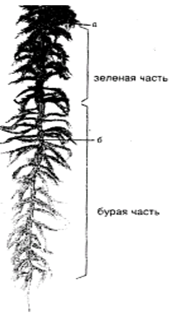
Рисунок 9 - Иллюстрация метода « индивидуальных меток».
а – место кольцевания, б- нахождение кольца через год роста, (а-б) – приросшая за год часть стебля
Приросшая часть стебля отрезается, высушивается до абсолютно сухого веса и взвешивается. Полученный вес соответствует годичному приросту одного растения. Каждый помеченный экземпляр отбирается в моновидовом сообществе с площадки размером 100 см2.
На этих же площадках определялось количество стеблей или плотность. Продукция каждого вида сфагновых мхов равняется произведению среднего прироста одного растения на количество растений на единице площади. Среднее из суммы приростов разных видов мхов из рода Sphagnum на единице площади является годичной продукцией мхов экосистемы (ANPM).
Для сообщества в целом общая надземная продукция представляет
ANP=ANPT + ANPk1 + ANPk2 + ANPM.
Все полученные данные обрабатывались статистическими методами.
4. ЗАПАСЫ И ДИНАМИКА ПРИРОСТА ФИТОМАССЫ МХОВ
Растительные сообщества выделенных экосистем по многим параметрам отличаются друг от друга: по водно-минеральному питанию, растительности, по запасам живой фито-массы, продукции, по приуроченности к различным элементам рельефа, по видовому разнообразию. Один из основных критериев, показывающий стабильность и устойчивость экосистемы, определяется видовым разнообразием.
Видовое разнообразие в рассмотренных экосистемах сильно варьирует, и прослеживается зависимость от положения в рельефе и водно-минерального питания экосистемы (таблица 1).
Таблица 1 -Распределение видов в различных экосистемах, количество видов
| Группа растений | Повышенный микрорельеф | Пониженный микрорельеф | |||||
| Экосистемы | Рям | Гряда | Мочажины | Топь | |||
Тип водно-минерального питания Подобные работы:
Актуально:
| |||||||