Параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданию
Глава 1. Обзор литературы
1.1 Системы транспорта калия в митохондриях
1.1.1 Транспорт калия в митохондрии
1.1.2 Молекулярные структуры, ответственные за транспорт калия в МХ
1.1.3 Физиологическое значение транспорта калия в МХ
1.1.4 АТФ-ингибируемые калий-транспортирующие каналы
1.2АТФ-зависимый калиевый канал цитоплазматической мембраны
1.2.1 Структурная организация цитоплазматического АТФ-зависимого калиевого канала
1.3 АТФ-чувствительный калиевый канал внутренней мембраны МХ
1.3.1 Структурная организация митоКАТФ канала
Глава 2. Модуляторы
2.1 Модуляторы митоКАТФ канала
2.1.1 Метаболические модуляторы митоКАТФ канала
2.2 Функциональная роль митоКАТФ
2.2.1 Активация митоКАТФ в развитии устойчивости организма к гипоксии
2.2.2 Механизмы защиты сердца при гипоксии, опосредованные активацией митоКАТФ
2.2.3 Феномен прерывистой гипобарической тренировки
2.3 Выделение МХ
2.3.1 Выделение МХ печени крысы
2.3.2 Выделение МХ сердца крысы
2.3.2 Выделение и очистка митоКАТФ канала
Глава 3. Изучение энергозависимого входа К+ в МХ методом спектрофотометрии
3.1 Изучение ДНФ-индуцированного выхода ионов калия из МХ
3.2 Получение и очистка антител к белку с молекулярной массой 55 кДа.
3.2.1 Подготовка белка с м.м. 55 кДа: выделение и очистка
3.3 Иммунизация и анализ препарата антител
3.3.1 Детекция специфических антител и определение титра
3.3.2 Вестерн-Блот анализ
3.4 Исследование ДНФ-индуцированного выхода К+ из митохондрий с помощью К+-селективного электрода
3.5 Реконструкция белка в БЛМ
3.6 Иммунноэлектронная микроскопия
3.7 MS-MALDI-TOF/TOF- анализ
3.8 Очистка антител к АТФ-зависимому белку с м.м. 55 кДа
3.9 Очистка антител к АТФ-зависимому белку с м.м. 55 кДа на колонке с иммобилизованным Белком А
3.10 Ингибиторный анализ с использованием антител к белку
с м.м. 55 кДа
Глава 4. Выделение комплекса цитоплазматических мембран и микросом печени крыс
4.1 Метод отбора высоко- и низкоустойчивых животных
Глава 5. Результаты и обсуждения
5.1 Параметры функционирования митоКАТФ канала у крыс с различной резистентностью, а также у животных, адаптированных к гипоксии
5.1.1 Изучение параметров дыхания и окислительного фосфорилирования в МХ печени и сердца крыс с различной резистентностью к гипоксии
5.1.2 Изучение параметров АТФ-зависимого транспорта К+ в МХ печени сердца крыс с различной резистентностью к гипоксии
5.1.2 Изучение параметров АТФ-зависимого транспорта К+ в МХ печени сердца крыс с различной резистентностью к гипоксии
5.2 Изучение структурной организации митохондриального АТФ-зависимого калиевого канала
5.2.1 Определение гомологии белка с м.м. 55 кДа методом MS-MALDI-TOF/TOF
5.3 Ингибиторный анализ активности митоКАТФ канала с использованием антител, полученных на белок с м.м. 55 кДа
5.3.1 Определение степени чистоты белка, используемого для иммунизации
5.4 Иммунизация и определение титра полученных антител
5.4.1 Определение специфичности полученных антител
5.4.2 Выделение иммуноглобулинов G (IgG) из антисыворотки и проведение ингибиторного анализа
5.5 Электронная микроскопия МитоКАТФ канала
ЗАКЛЮЧЕНИЕ
ВЫВОДЫ
ЛИТЕРАТУРА
СПИСОК СОКРАЩЕНИЙ
МитоКАТФ – митохондриальный АТФ-чувствительный калиевый канал
ЦитоКАТФ – цитоплазматический АТФ-чувствительный калиевый канал
5-ГД – 5-гидроксидекановая кислота
Глиб – глибенкламид
АТФ – аденозин-5’-трифосфат
УТФ – уридин-5’-трифосфат
УДФ – уридин-5’-дифосфат
УМФ – уридин-5’-монофосфат
ГТФ – гуанин-5’-трифосфат
МХ – митохондрии
АТ – антитела
ФН – неорганический фосфат
АФК – активные формы кислорода
KIR – inward rectifying K+ channels (канальная субъединица митоКАТФ канала)
SUR – sulphonyl urea receptor (регуляторная субъединица митоКАТФ канала,
чувствительная к сульфонилмочевинам)
PCO’s – potassium channel openers (активаторы калиевых каналов)
NFBs – nucleotide phosphate binding domains (нуклеотидсвязывающие участки)
БЛМ – бислойные липидные мембраны
ФИА - феномен ишемической адаптации
ГТ – гипоксическая тренировка
НУ – крысы, низкоустойчивые к гипоксии
ВУ – крысы, высокоусточивые к гипоксии
Введение
Митохондриальный АТФ-ингибируемый калиевый канал (митоКАТФ), осуществляющий вход калия в МХ, был обнаружен методом пэтч-кламп во внутренней мембране МХ в 1991 г. (Inoue et al., 1991). Однако, еще в 1981 г. в лаборатории проф. Мироновой был выделен белок с м.м. 55 кДа, обладающий свойствами данного канала. (Миронова и др., 1981). Позднее было показано, что выделенный белок-канал ингибируется физиологическими концентрациями АТФ (Paucek et al., 1992; Миронова и др., 1996 (I)).
В настоящее время достаточно хорошо исследованы биофизические свойства митохондриального калиевого канала и его физиологическая роль (Миронова и др., 1996 (I, II); Paucek et al., 1992; Inoue et al., 1991; Garlid et al., 1997; Mironova et al., 1999; 2004). Интерес к исследованию этого канала в последнее время возрос, поскольку было показано, что он, а именно его активация, играет ключевую роль в защите миокарда при ишемии (Grover et al., 1992; Garlid et al., 1997; Vanden Hoek, 2000). Найден целый ряд синтетических активаторов митоКАТФ, являющихся потенциальными кардиопротекторами (Gross et al., 1992; Liu et al., 1998; Sato et al., 1998; Tsai et al., 2002). Недавно в лаборатории проф. Мироновой был обнаружен эффективный природный метаболический активатор митоКАТФ – уридин-5’-дифосфат (УДФ) (Mironova et al., 2004; Негода А.Е., 2004). Метаболические активаторы канала имеют ряд преимуществ, по сравнению с их синтетическими аналогами, поскольку их концентрацию в клетке можно регулировать и они не обладают отрицательными побочными эффектами. Действие УДФ, как активатора К+-канала, и, следовательно, кардиопротектора, до настоящего времени не было изучено на животных.
Помимо важной роли митоКАТФ в защите миокарда от ишемических повреждений, некоторые исследователи предполагают участие активации канала в формировании устойчивости организма к кислородному голоданию (Zhu et al., 2003). Однако прямые доказательства участия канала в адаптации к гипоксии до настоящего времени не получены, то есть, данный феномен также требует дополнительных исследований.
К настоящему времени нами разработана рабочая модель структуры и регуляции канала, в соответствии с которой митоКАТФ канал состоит из двух субъединиц – канальной (с молекулярной массой 55 кДа) и регуляторной (с молекулярной массой 63 кДа). Однако вопрос о структуре канала и возможной гомологии с другими белками пока остается открытым. Изучение структуры митоКАТФ позволит исследовать функцию и регуляцию канала на молекулярном уровне.
В связи с этим, целью данной работыбыло: изучить структурную организацию митоКАТФ канала, кардиопротекторное действие УДФ при ишемии миокарда, исследовать параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданию.
Таким образом, в работе были поставлены следующие задачи:
1) Исследовать параметры функционирования митоКАТФ у крыс с различной устойчивостью к гипоксии, а также у животных, адаптированных к недостатку кислорода.
2) Определить гомологию структуры исследуемого белка с м.м. 55 кДа аминокислотным последовательностям известных белков.
3) Получить специфические поликлональные антитела на белок-канал с м.м. 55 кДа, формирующий при встраивании в искусственные мембраны АТФ-ингибируемые К+ каналы.
4) Провести ингибиторный анализ АТФ-чувствительного транспорта калия в нативных МХ с использованием полученных антител (АТ) с целью доказательства принадлежности белка с м.м. 55 кДа к системе АТФ-зависимого транспорта К+ в МХ.
5) Исследовать иммунолокализацию МитоКАТФ канала на срезе гепатоцитов и кардиомиоцитах при помощи электронной микроскопии
Глава 1. Обзор литературы1.2 Системы транспорта калия в митохондриях
1.1.1 Транспорт калия в митохондрии
Транспорт К+ в митохондриях (МХ) имеет большое функциональное значение, так как концентрация К+ в цитоплазме значительно превышает концентрацию других катионов, и появление любой неконтролируемой проницаемости митохондриальной мембраны для К+ может представлять угрозу осмотической целостности и функциональной интегральности МХ (Brierley et al., 1983). МХ занимают 40% внутриклеточного пространства в сердечной клетке и до 20% - в клетке печени и по данным (Rottenberg, 1973), в свободном состоянии в матриксе МХ содержится 95 нмоль К+/мг белка МХ, в слабосвязанном состоянии – 45 нмоль и в прочносвязанном – 5 нмоль. Концентрация свободного калия в цитоплазме составляет 90-120 нмоль.
Хемиосмотическая теория Митчела включает четыре постулата, последние два из которых имеют отношение к системе транспорта катионов. Согласно этим двум постулатам, внутренняя мембрана МХ: а) непроницаема для Н+, ОН- и всех прочих ионов; б) содержит ряд белков-переносчиков, осуществляющих транспорт неорганических ионов и необходимых метаболитов. Таким образом, по Митчелу, внутренняя мембрана МХ непроницаема для катионов и анионов. Однако из-за большого электрохимического потенциала (Δφ) (~ –200 мВ с матриксной стороны) одновалентные катионы (К+ и Na+) могут диффундировать через липидный бислой. Проблема такой утечки будет наиболее актуальна для ионов К+, как основного катиона цитозоля и матрикса, хотя скорость такой диффузии будет не велика.
Что касается специфических систем транспорта, то было установлено, что в МХ существуют системы: 1) электрогенного входа калия и 2) К+/Н+-антипортер (Chavez et al., 1977; Diwan, 1981; Garlid, 1980), о существовании которого говорил Митчел (Mitchell and Moyle, 1969).
В работах с использованием радиоактивного калия (42К+) было показано, что активный транспорт К+ в МХ осуществляется электрогенно и что существует специфическая К+-транспортирующая система, функционирующая подобно ионофорам, катализирующим унипорт калия (Gamble, 1957, 1962; Judan et al., 1965; Rottenberg, 1973; Chavez et al., 1977).
В качестве системы электрогенного транспорта К+ в МХ предлагались следующие варианты: 1) подвижный переносчик или канал (Jonston and Griddle, 1978; Panov et al., 1980; Jung et al., 1982); 2) электродиффузное движение через гидрофильные поры и 3) вход калия с участием белковых транспортных систем, которые помимо своей основной функции, могут осуществлять транспорт К+ (Brierly, 1980, 1983).
1.1.3 Молекулярные структуры, ответственные за транспорт калия в МХ
В настоящее время в литературе в качестве транспортных систем, осуществляющих электрогенный транспорт калия в МХ, рассматриваются следующие структуры: 1) АТФ/АДФ-антипортер или аденинуклеотидтранслоказа (АНТ) (Panov et al., 1980; Le Quoc et al., 1988) и 2) специфические белки, которые образуют во внутренней мембране МХ канал для электрогенного входа ионов калия (Миронова и др., 1981; Diwan et al., 1988; Paucek et a., 1992). В первом случае предполагается, что АНТ, помимо основных функций, может работать как система электрофоретического транспорта ионов калия в МХ при действии на МХ субмикромолярных концентраций ионов кальция, и как неспецифическая пора для низкомолекулярных соединений при повреждении МХ высокими концентрациями Ca2+ (Jung and Brierley, 1981; 1982; Halestrap et al., 1986).
Что касается специфических белков, следует отметить, что в лаборатории Мироновой в 1981 г. методом водно-этанольной экстракции из МХ сердца быка был выделен и очищен белок с м.м. 55 кДа, который при реконструкции в бислойные липидные мембраны (БЛМ) образовывал К+-селективные каналы проводимости (Миронова и др., 1981), которые, как было показано позднее, ингибируются физиологической концентрацией АТФ и глибенкламидом, и относятся к семейству АТФ-чувствительных калиевых каналов (Миронова и др., 1996 (I); Mironova et al., 1999). Структура митохондриального КАТФ канала не определена (Paucek et al., 1992).
Диван с соавторами, используя детергент Тритон Х-100, из МХ мембраны печени крыс выделили белок с м.м. 53 кДа (Diwan et al., 1988). Реконструировав его в липосомы, авторы показали, что этот белок также обладает свойствами, характерными для К+ унипортера.
Известно, что в МХ помимо системы электрогеннного входа К+ существует система электронейтрального выхода К+ в обмен на Н+ (Garlid and Paucek, 2003). При этом внешняя МХ мембрана не препятствует дальнейшему обмену небольшими ионами с цитоплазмой.
Система электрогенного входа калия и электронейтрального К+/Н+-обменника образуют К+ цикл МХ (Рис.1).
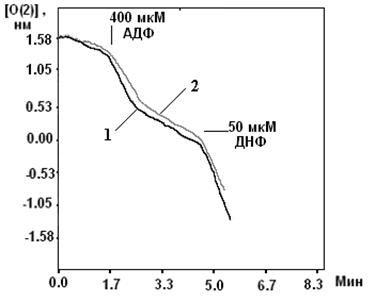
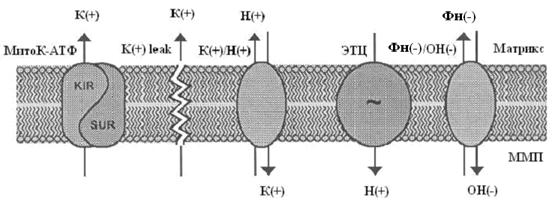
Рис. 1. МХ калиевый цикл по Garlid and Paucek, 2003. ЭТЦ – электрон-транспортная цепь; ММП – межмембранное пространство; Фн – неорганический фосфат.
Электрогенный выброс протонов электрон-транспортной системой генерирует мембранный потенциал, который, в свою очередь, стимулирует диффузию К+ в матрикс МХ («утечка калия» или К+ leak) и вход иона, опосредованный неким специфическим калиевым каналом. Такой обмен Н+ на К+ подщелачивает матрикс, вызывая вход фосфата по электронейтральному Фн-Н+ симпортеру (Фн – неорганический фосфат) (Рис.1).
Вход К+ сопровождается накоплением осмотически облигатной воды, что проявляется в набухании МХ. Излишнее их набухание может угрожать целостности этих органелл, следовательно, избыток К+ необходимо удалять. Эту функцию выполняет белок с м.м. 82 кДа - К+/Н+-антипортер (Brierley G., 1976; Garlid K., et al., 1980; 1988). Следует отметить, что вход К+ путем диффузии слишком мал, чтобы существенно влиять на изменение объема матрикса МХ. Интенсивный вход калия в МХ обеспечивается специфическими белками, осуществляющими электрофоретический вход К+ в МХ (Mironova G., 1981; Diwan J., et al., 1988; Paucek et al., 1992).
1.1.3 Физиологическое значение транспорта калия в МХ
Предполагается, что основное физиологическое значение систем транспорта К+ в МХ связано с регуляцией объема МХ матрикса. Объем МХ определяется калиевыми токами через внутреннюю мембрану. Когда вход и выход К+ находятся в равновесии, калиевые потоки определяются электронейтральным током анионов и осмотически облигатной воды (Garlid, 1988). Так как концентрация ионов калия в матриксе и в цитоплазме практически одинаковая, транспорт калия мало влияет на матриксную концентрацию калия, но может иметь большое влияние на объем МХ. Небольшое нескомпенсированное увеличение входа К+ в МХ может удваивать их объем в течение 1-2 минут (Garlid, 1979). В свою очередь, увеличение матриксного объема стимулирует активность дыхательной цепи, что было показано на МХ сердца и печени (Nichols and Lindberg, 1972; Halestrap, 1989). Окисление жирных кислот также чувствительно к изменению объема матрикса (Halestrap, 1987).
Увеличение в МХ концентрации К+ и последующее изменение их объема имеет значение и в регуляции митохондриальных процессов у зимоспящих животных (Fedotcheva et al., 1985; Бакеева и Брустовецкий, 1993), а также при адаптации животных к холоду (Nedergraard and Cannon, 1987). В ряде работ установлено, что in vivo, гормон, участвующий в регуляции теплопродукции (тироксин), активирует системы электрогенного транспорта К+, вызывая низкоамплитудное набухание МХ (Halestrap, 1987; Shears an Brouk, 1980). В нашей лаборатории, было показано, что активность системы электрогенного транспорта ионов калия прямо коррелирует с интенсивностью термогенеза (Федотчева и др., 1984; Миронова и др., 1986), что непосредственно связано с увеличением содержания ионов калия в МХ бурой жировой ткани и печени (Скарга, 1994). Активацию транспорта калия при выходе животного из спячки, связывают с активацией футильного цикла К+ в МХ, приводящей к увеличению теплопродукции, до начала синтеза АТФ в МХ (Миронова и др., 1986).
Обнаружено также, что митоКАТФ канал играет существенную роль в нормальной физиологии миокарда, регулируя объем митохондрий и продукцию активных форм кислорода (АФК) (Garlid et al., 2003a).
В последнее время интерес ученых к митоКАТФ каналу вызван обнаружением его роли в защите сердца от инфаркта. Исследования были стимулированы открытием феномена кардиопротекторного действия прерывистой гипоксии, приводящей к активации митоКАТФ (Murry et al., 1986). Установлено, что фармакологические активаторы митоКАТФ канала предохраняют сердце от ишемических повреждений (Garlid et al., 1997).
1.1.4 АТФ-ингибируемые калий-транспортирующие каналы
К+ каналы, активность которых ингибируется физиологическими концентрациями АТФ впервые были обнаружены в цитоплазматической мембране кардиомиоцитов (Noma, 1983). С тех пор, аналогичные каналы были найдены в β клетках поджелудочной железы (Dunne and Petersen, 1991), в скелетной мускулатуре (Spruce et al., 1985; Woll et al., 1989), в нервных клетках (Jonas et al., 1990) и гладких мышцах (Standen et al., 1989). Активность АТФ-чувствительных калиевых каналов (КАТФ каналов) связана с уровнем биоэнергетического метаболизма клетки (концентрацией АТФ) и электрическими свойствами возбудимости плазматической мембраны. Микромолярные концентрации АТФ и сульфонилмочевины – класс гипогликемических соединений, использующихся при лечении диабета, регулируют активность всех представителей семейства КАТФ каналов (Ashcroft et al., 1989). Концентрация кальция и изменение мембранного потенциала практически не влияют на активность этих каналов (Ashcroft and Ashcroft, 1990). В то же время, в кортикальных и гипоталамических нейронах КАТФ каналы (Ashford et al., 1989; Ashford et al., 1988) менее чувствительны к АТФ (для их ингибирования нужны миллимолярные концентрации). В эпителиальных клетках (назальные полипы) также были обнаружены КАТФ каналы с низкой чувствительностью к АТФ, но активирующиеся микромолярными концентрациями Ca2+ (Kunzelmann et al., 1989). При этом, все вышеперечисленные каналы высокоселективны для ионов К+.
В МХ мембране также были обнаружены АТФ-зависимые К+ каналы. Так, в 1991 г. впервые с использованием метода петч-кламп (Inoue et al., 1991) на митопластах было показано, что во внутренней мембране МХ присутствуют высокоселективные по К+ каналы с невысокой проводимостью. Обнаружено, что каналы могут обратимо ингибироваться АТФ, а так же 4-аминопиридином и специфическим ингибитором цитоплазматического КАТФ канала – глибенкламидом с матриксной стороны (Inoue et al., 1991). Однако, как было сказано выше, еще в 1981 году в лаборатории проф. Мироновой во внутренней мембране МХ был обнаружен К+-селективный АТФ-ингибируемый канал (Миронова и др., 1981; 1996; 1997; 1999), принадлежащий к семейству КАТФ каналов (Paucek et al., 1992). Данный канал имеет те же характеристики проводимости, что и канал, обнаруженный методом пэтч-кламп. В настоящее время он интенсивно изучается, поскольку играет важную роль в функционировании клетки. Однако вопрос о его структуре требует дальнейших исследований.
1.2 АТФ-зависимый калиевый канал цитоплазматической мембраны
К настоящему времени КАТФ каналах плазматической мембраны хорошо изучены. Изначально их классификация велась по величине проводимости одиночных каналов (Ashcroft and Ashcroft, 1990), а позднее, с развитием молекулярно-биологических методик, по аминокислотному составу белковой молекулы канала (Yokoshiki et al., 1998).
1.2.1 Структурная организация цитоплазматического АТФ-зависимого калиевого канала
Молекулярно-биологические и электрофизиологические исследования последних лет позволили определить структуру цитоКАТФ канала. Исследуемый канал состоит из двух белков: KIR – inward rectifying K+ channels, формирующий пору канала, и SUR – sulphonyl urea receptor, регуляторная субъединица, придающая каналу чувствительность к модуляторам. SUR-субъединица содержит нуклеотидсвязывающие участки, локализующиеся на цитоплазматической стороне, а также участки связывающие активаторы калиевых каналов (potassium channel activators, PCOs) и MgАДФ. Каналы подсемейства KIR6.0 сами обладают чувствительностью к АТФ. Роль SUR заключается, вероятно, в облегчении доступа АТФ к АТФ-связывающему участку KIR (Tucker et al., 1997; Yokoshiki et al., 1998). В настоящее время механизмы молекулярного взаимодействия KIR и SUR окончательно не выяснены.
Результаты исследования электрофизиологических свойств нативных КАТФ каналов позволяют предположить, что они принадлежат к классу калиевых каналов внутреннего выпрямления (KIR). В 1993 году на многие KIR каналы (KIR1.1 (Ho et al., 1993), KIR2.1 (Kubo et al., 1993), KIR3.1 (Dascal et al.,1993)) были получены клоны. Их выделили в отдельное подсемейство KIR-каналов (KIR1.0-6.0), основанное на унифицированной номенклатуре (Chandy and Gutman, 1993). Гидрофобные участки клонированных KIR каналов содержат два мембранопересекающих сегмента М1 и М2, образующих пору. Подобно потенциалзависимым К+ каналам, KIR каналы, вероятно, формируют тетрамер (Doupnik et al., 1995; Salkoff and Jegla, 1995; Nichols et al., 1996; Nichols and Lopatin, 1997).
Также был клоирован регуляторный белок, содержащий участок, связывающий сульфонилочевину (SU-связывающий сайт) (Aguillar-Bryan et al., 1995; Thomas et al., 1995; Inagaki et al., 1996; Isomoto et al., 1996). Клонированные регуляторные белки получили название SUR. SUR содержит два нуклеотидсвязывающих участка (NFBs) с цитоплазматической стороны. N-конец находится с внешней стороны мембраны и содержит два домена, предположительно из 9 и 4 пронизывающих мембрану спиралей (Рис.2) (Yokoshiki et al., 1998).
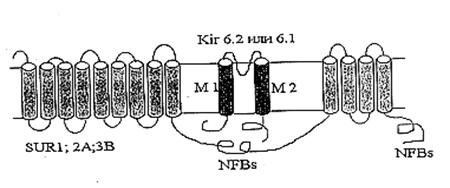
Рис. 2. Схематическое изображение цитоплазматического КАТФ канала как комплекса KIR и SUR (YokoshikiH. etal., 1998)
Считается, что нативный КАТФ канал представляет собой комплекс из четырех KIR6.x и четырех SUR субъединиц (Clement et al., 1997). Различные типы регуляторных субъединиц, SUR1, SUR2A и SUR2B, обнаружены, соответственно, в поджелудочной железе, сердце и гладких мышцах (Ammala et al., 1996; Inagaki et al., 1996; Yamada et al., 1997).
1.3 АТФ-чувствительный калиевый канал внутренней мембраны МХ
В 1981 г. из внутренней мембраны МХ был выделен К+-селективный канал (митоКАТФ) (Миронова и др., 1981), который ингибируется физиологическими концентрациями АТФ (Paucek et al., 1992; Миронова и др., 1996 (I)). В 1991 г. этот канал был впервые обнаружен во внутренней мембране МХ методом пэтч-кламп (Inoue et al., 1991), позднее появились аналогичные работы других исследователей (Nakae et al., 2003; Fikret et al., 2004; Dahlem et al., 2004; Bednarczyk et al., 2005). В настоящее время канал активно изучается, поскольку обнаружено, что он участвует в адаптации организма к гипоксии (Garlid et al., 1997). В настоящее время основными методами выделения АТФ-ингибируемого МХ калиевого канала (митоКАТФ) является метод водно-этанольной экстракции (Миронова и др., 1981; Миронова и др., 1996 (I)) и солюбилизация белка-канала из мембраны Тритоном Х-100 с последующей очисткой ионообменной хроматографией. При этом митоКАТФ канал выделяют в виде белка с молекулярной массой 55 кДа (Mironova et al., 2004).
1.3.1 Структурная организация митоКАТФ канала
Так как митоКАТФ имеет ряд общих свойств с цитоКАТФ и регулируется общими регуляторами, было предположено, что по структуре он близок к цитоплазматическому каналу (Mironova et al., 1999) и состоит из канальной субъединицы (Mironova et al., 2004) и регуляторной (Bajgar et al., 2001). Белок с м. м. 55 кДа, выделенный из внутренней мембраны МХ печени крысы, является, по-видимому, канальной субъединицей митоКАТФ. При встраивании в БЛМ этот белок формирует АТФ-зависимые каналы (митоKIR). Активность этих каналов не блокируется глибенкламидом и специфическим ингибитором митоКАТФ 5-ГД, и не активируется кромакалимом и ддиазоксидом. Однако эти препараты влияют на АТФ-зависимый калиевый транспорт в изолированных МХ, где есть обе субъединицы канала (Mironova et al., 2004).
МитоКАТФ, также как и клеточный, является, по всей вероятности, гетеромультимером, состоящим из калиевого канала, белка с молекулярной масой 55 кДа, который имеет выпрямляющие свойства и который, по аналогии с цитоплазматическим каналом, был назван митоKIR (от англ. Inward rectifying) (Mironova et al., 2004) и рецептора, чувствительного к сульфонилмочевинам и поэтому названного митоSUR (от англ. sulphonylurea) (Рис. 3) (Mironova et al., 1999; Bajgar et al., 2001).
Кi для АТФ на реконструированном в БЛМ митоKIR выше, чем в интактных МХ и Mg2+ для этого ингибирования не требуется. Тогда как для АТФ-ингибирования канала, локализованного в интактных МХ, присутствие магния было необходимо (Mironova et al., 2004). Те же различия были обнаружены при сравнительном изучении влияния АТФ и сульфонилмочевин на цитоKIR и целый цитоКАТФ (Tucker et al., 1997). Известно, что основной участок связывания АТФ в цитоКАТФ локализован на канальной субъединице. Регуляторная субъединица повышает сродство канальной субъединицы к АТФ и обеспечивает чувствительность целого канала к активаторам и ингибиторам (Tucker et al., 1997).
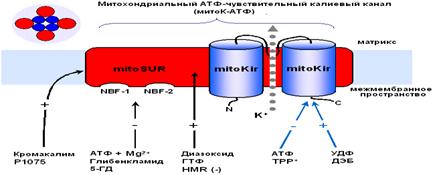
Рис.3. Рабочая модель структуры МХ АТФ-зависимого К+ канала (Mironova et al., 2004)
Приведенные выше результаты исследований митоКАТФ подтверждают предположение о том, что белок с м.м. 55 кДа является канальной субъединицей целого митоКАТФ. Функцию регуляторной субъединицы выполняет, вероятно, белок с м.м. 63 кДа, связывающийся с меченым глибенкламидом (Bajgar et al., 2001).
В то же время, результаты исследований, проведенных на МХ сердца, с использованием антител на Kir6.1 и Kir6.2, а также на SUR1 и SUR2, показали, что в МХ кардиомиоцитов присутствуют Kir6.1 и Kir6.2, но нет ни SUR1, ни SUR2. Однако в них обнаруживается SUR2-подобный белок, который, возможно, является сплайсированным вариантом SUR2 (Lacza et al., 2003 (I)).
При исследовании митоКАТФ мозга, близкого по структуре к цитоКАТФ (Lacza et al., 2003 (II)), с использованием АТ к KIR6.1 и KIR6.2, а также SUR1 и SUR2 субъединицам цитоКАТФ было показано, что очищенные МХ мозга содержат преимущественно KIR6.1 и, в меньшей степени, KIR6.2 и не содержат SUR1 и SUR2. Согласно выводам авторов, в формировании митоКАТФ мозга участвует укороченный вариант SUR2 или белок, подобный по структуре SUR2. Кроме того, показано, что МХ мозга содержат в 8 раз больше KIR6.1 и SUR2-подобного белка, по сравнению с общим белком ткани мозга, что подтверждает локализацию этих белков в МХ (Lacza et al., 2003 (II)). Тот факт, что м.м. митоKIR мозга несколько ниже (50 кДа), чем у митоKIR печени (Mironova et al., 1999; 2004), предполагает наличие тканевой специфичности в структуре каналов.
Было показано, что при встраивании в липосомы или липидные бислойные мембраны комплекса митохондриальных белков, состоящего из АТФ связывающего кассетного белка-1 (mАВС1 от англ. Mitochondrial binding casset), АТФ-азы, сукцинатдегидрогеназы, аденилаттранслоказы и фосфатного переносчика, в мембране образуются чувствительные к АТФ и глибенкламиду каналы (Ardehali et al., 2004). Это послужило основой для другого варианта модели субъединичной организации митоКАТФ (Ardehali and O'Rourke, 2005). Однако нельзя исключить наличие в этом белковом комплексе других митохондриальных белков, включая митоKIR, тем более что белок с м.м. 55 кДа в нём присутствует. Поcкольку этот мультикомплекс в липосомах индуцировал открытие неспецифической Са2+-зависимой поры в присутствие ионов Са2+ (Ardehali et al., 2004), это говорит о возможном загрязнении исследуемого комплекса порином и другими белками. Было показано, что один из белков этого комплекса, а именно mАВС1, играет важную роль в защите клеток миокарда от окислительного стресса (Ardehali et al., 2005). Однако кардиопротекторное действие этого белка не снималось ингибиторами митоКАТФ - глибенкламидом и 5-НD. Поэтому, по мнению авторов, пока не получено достаточно доказательств для утверждения того, что mАВС1, имеющий, также как и митоКIR, м.м. 55 кДа, является частью митоКАТФ.
Таким образом, вопрос структуры митоКАТФ по-прежнему остается открытым. В связи с чем, в представленной работе была предпринята попытка определить относится ли белок с м.м. 55 кДа, выделяемый из внутренней мембраны МХ методом водно-этанольной экстракции, к системе АТФ-зависимого транспорта К+. Также, был проведен анализ гомологии структуры исследуемого белка последовательностям известных белков.
Глава 2. Модуляторы
2.1 Модуляторы митоКАТФ канала
2.1.1 Метаболические модуляторы митоКАТФ канала
В связи с выраженным кардиопротекторным действием активаторов митоКАТФ в настоящее время во многих лабораториях ведется поиск новых синтетических активаторов канала, которые могут быть потенциальными кардиопротекторами (Ockaili et al., 2002; Kopustinskene et al., 2001). В нашей лаборатории помимо синтетического донора электронов ДЕБ, который селективно активировал митоКАТФ (Mironova et al., 1999; Mironova et al., 2004), был обнаружен ряд природных, то есть метаболических активаторов этого канала.
Дифосфонуклеотиды
Показано, что дифосфонуклеотиды (АДФ и ГДФ) активируют реконструированную в БЛМ канальную субъединицу (Рис. 2) (Grigoriev et al., 1999; Mironova et al., 1999). Данные по активирующему действию ГДФ в дальнейшем были подтверждены и другими исследователями (Bednarczyk et al., 2005). Недавно в нашей лаборатории было показано, что в ряду дифосфонуклеотидов наиболее эффективным является уридиндифосфат (УДФ) (Mironova et al., 2004).
В МХ УДФ (~20 мкМ) активирует митоКАТФ, и эта активация снимается глибенкламидом и 5-ГД (Рис. 4) (Mironova et al., 2004). Концентрации УДФ, неоходимые для активации реконструированного в БЛМ митоKIR и целого канала в нативных МХ, одинаковы. Следовательно, участок связывания УДФ локализуется на канальной субъединице. Для активации цитоКАТФ нужны значительно большие концентрации этого препарата (Alekseev et al., 1998).
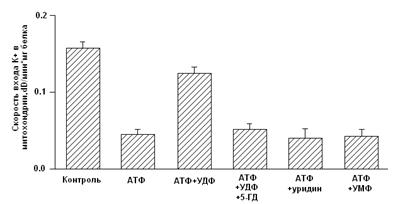
Рисунок 4. Активация УДФ целого митоКАТФ канала в нативных МХ (Mironova et al., 2004)
Ранее на моделях тотальной и регионарной ишемии изолированных перфузируемых сердец крыс было показано, что уридин и УМФ проявляют антиаритмическую активность в отношении реперфузионных аритмий, предупреждают развитие миокардиального станинга при постишемической реперфузии, препятствуя депрессии сократительной функции миокарда и предотвращая развитие контрактуры (Eliseev et al., 1989).
В представленной работе была предпринята попытка определить, обладают ли уридиновые нуклеотиды кардиопротекторным действием и связано ли их влияние с активацией цито- и митоКАТФ каналов на модели искусственного острого инфаркта миокарда.
2.2 Функциональная роль митоКАТФ
Являясь одним из компонентов калиевого цикла МХ, митоКАТФ участвует в поддержании объема МХ матрикса и мембранного потенциала (Garlid, 1998). В лаборатории проф. Мироновой показано, что активация митоКАТФ играет важную роль при выходе животных из состояния зимней спячки (Fedotcheva et al., 1985; Миронова и др., 1986). МитоКАТФ участвует также в ингибировании процесса апоптоза клетки, блокируя высвобождение цитохрома С, предотвращая снижение мембранного потенциала и чрезмерное увеличение концентрации Ca2+, сохраняя, таким образом, функции МХ, что было показано на b-клетках поджелудочной железы (Akao et al., 2001), а затем и на церебральных нейронах (Teshima et al., 2003). Кроме того, митоКАТФ является посредником в процессе регуляции транскрипции некоторых генов. Его активация вызывает небольшое увеличение продукции активных форм кислорода (АФК) (Chandel et al., 1998).
При патологии активация митоКАТФ частично востанавливает мембранный потенциал, стимулируя дальнейший перенос протонов, ингибируя накопление внутримитохондриального кальция и предотвращая распад энергетических фосфатов путем создания оптимального для синтеза АТФ электрохимического градиента (Szewczyk, 1996; Xu et al., 2001). Активация митоКАТФ способствует разобщению электронного транспорта, ускоряя, таким образом, дыхание, что приводит к окислению проксимальных переносчиков электронов дыхательной цепи (Liu et al., 1998). Согласно данным Гарлида, активация канала приводит к набуханию МХ, что способствует поддержанию архитектуры внутренней мембраны и сохранению активности дыхательной цепи. Это позволяет поддерживать уровень клеточного АТФ и, таким образом, способствует восстановлению функций миокарда во время реперфузии (Garlid, 2000). Как было показано ранее, набухание матрикса увеличивает скорость окислительного метаболизма, активирует как окисление жирных кислот, так и дыхание и синтез АТФ (Halestrap, 1989).
2.2.1 Активация митоКАТФ в развитии устойчивости организма к гипоксии
В 1983 г. был открыт феномен прерывистой гипоксии, важную роль в котором играет активация митоКАТФ (Murry et al., 1983; Liu et al., 1998).
Феномен прерывистой гипоксии был назван «preconditioning» или «ишемической толерантностью» (Murry et al., 1986; Kitigawa, 1990). Позднее он получил название «феномен ишемической адаптации» (ФИА) (Yellon et al., 1998; Петрищев и др., 2001), и в настоящее время признан наиболее эффективным способом уменьшения размера инфаркта миокарда (Kloner et al., 1998). Справедливости ради надо отметить, что адаптация, создаваемая кратковременной гипоксией, была обнаружена ранее (Poupa et al.,1966; Meerson et al., 1973).
Клеточные механизмы ФИА в настоящее время широко изучаются. Считается, что защитный эффект является трехступенчатым и включает в себя последовательные триггерные и конечные эффекторные механизмы, причем наиболее вероятным эффектором является митоКАТФ (Garlid et al., 1997; Yellon et al., 1998; Grover and Garlid., 2000; Петрищев и др., 2001).
ФИА развивается в виде двух волн: первая волна проявляется в течение нескольких часов, затем исчезает и снова появляется в виде второй волны (Yellon et al., 1998), которая длится в течение недели, а иногда и месяца (Carroll et al., 2000). Установлено, что во второй стадии адаптации к гипоксии также участвует митоКАТФ (Wang et al., 2001). В развитии толерантности к ишемии мозга (Lacza et al., 2003), и в дистанционной ишемической адаптации сердца к гипоксии при создании коротких эпизодов окклюзии почечной артерии также задействован митоКАТФ (Pell et al., 1998).
Установлено, что специфические модуляторы митоКАТФ (см. таблицу 1) влияют на ФИА. Так активатор митоКАТФ, диазоксид, и неселективный активатор КАТФ каналов, кромакалим, имитируют «preconditioning», увеличивая время сокращения миокарда и способствовуя восстановлению функций изолированных сердец крыс, подвергавшихся общей ишемии миокарда с последующей реперфузией. Кi диазоксида для митоКАТФ (0.8 мкМ в МХ сердца быка) в 1000 раз превышала Кi для цитоКАТФ (Gross et al., 1992). Кромакалим проявлял аналогичный эффект в тех же концентрациях. Глибенкламид и 5-ГД блокировали действие диазоксида и кромакалима, следовательно, эффект активаторов на ФИА опосредован через КАТФ каналы (Auchampach, 1992; Garlid, 1997; Tsai et al., 1998; Vanden Hoek, 2000). Активирующе действие диазоксида на митоКАТФ показано также на изолированных МХ сердца крысы, что доказывает роль МХ канала в кардиопротекции (Gross et al., 1992). Полученные данные подтверждают также результаты исследований, по изучению флуоресценции флавопротеинов на изолированных кардиомиоцитах желудочков сердца кроликов (Liu et al., 1998; Sato et al., 1998). Однако поскольку было обнаружено, что диазоксид снижает мембранный потенциал на МХ, независимо от активации митоКАТФ канала, его кардиопротекторный эффект вызывает сомнение.
Показано, что потенциальный медиатор сигнального пути, ответственного за формирование ишемической адаптации, протеинкиназа С, (Sato et al., 1998; Liu et al., 1994; Sasaki et al., 2000) модулирует активность митоКАТФ. 5-ГД полностью блокирует этот кардиопротекторный эффект, подтверждая участие митоКАТФ в защите миокарда от ишемических повреждений. Следует отметить, что сами по себе глибенкламид и 5-ГД не оказывают никакого влияния (Gross and Fryer, 1999). Неспособность специфического ингибитора цитоКАТФ, HMR-1883, ингибировать анти-ишемический эффект активаторов митоКАТФ (Fryer et al., 2000), также говорит в пользу участия в защитном механизме МХ канала.
Следует отметить, что неселективные активаторы КАТФ каналов, такие как пинацидил, оказывая кардиопротекторное действие, вызывали побочный проаритмический эффект и приводили к существенному понижению давления у подопытных животных (Grover, 1997). Подобные нежелательные последствия способны свести на нет кардиопротекторное действие данных препаратов. Использов