Пластиды и их пигменты. Выделительные системы растений
Уральская государственная академия ветеринарной медициныКонтрольная работа
Выполнил:студент 1 курсазаочного факультетаотделениетехнологии ППСХПАвдеев Алексей ИвановичТроицк, 2006 год
ОГЛАВЛЕНИЕ
I. ПЛАСТИДЫ И ИХ ПИГМЕНТЫ, ФОТОСИНТЕЗ, НЕОБХОДИМЫЕ ДЛЯ НЕГО УСЛОВИЯ, ДЕЛЕНИЕ КЛЕТКИ
I.1 Пластиды и их пигменты
I.2 Фотосинтез, необходимые для него условия
I.3 Деление клетки
II. ВЫДЕЛИТЕЛЬНЫЕ СИСТЕМЫ РАСТЕНИЙ (железистые волоски, выделительные ходы, млечники и т.д.). ОХАРАКТЕРИЗУЙТЕ БАЛЬЗАМЫ, ЭФИРНЫЕ МАСЛА, СМОЛЫ, КАМЕДИ
II.1 Выделительные системы растений (железистые волоски, выделительные ходы, млечники и т.д.)
II.2 Характеристика бальзамов, эфирных масел, смол, камеди
III. ПОЧКИ, ТИПЫ ПОЧЕК ПО ПРОИСХОЖДЕНИЮ. ПРИВЕДИТЕ РИСУНОК ВЕГЕТАТИВНОЙ ПОЧКИ В ПРОДОЛЬНОМ РАЗРЕЗЕ IV. В КРУПНОМ МАСШТАБЕ НАРИСУЙТЕ СЕМЯЗАЧАТОК С ЗАРОДЫШЕВЫМ МЕШКОМ В ПРОДОЛЬНОМ РАЗРЕЗЕ. ИЗ КАКИХ ЧАСТЕЙ СЕМЯЗАЧАТКА РАЗВИВАЮТСЯ ТЕ ИЛИ ИНЫЕ ЧАСТИ СЕМЕНИ
V. СПОСОБЫ РАСПРОСТРАНЕНИЯ ПЛОДОВ И СЕМЯН. ПРИВЕДИТЕ ПРИМЕРЫ. БИОЛОГОИЧЕСКАЯ РОЛЬ РАСПРОСТРАНЕНИЯ ПЛОДОВ И СЕМЯН
VI. ПОНЯТИЕ О ВИДЕ РАСТЕНИЙ. СОСТАВЛЕНИЕ ВИДОВЫХ НАЗВАНИЙ РАСТЕНИЙ СОГЛАСНО БИНАРНОМУ МЕТОДУ К. ЛИННЕЯ. ВЫПИШЕТЕ ИЗ «СПИСКА ОСНОВНЫХ СЕМЕЙСТВ И ВИДОВ» 6 ВИДОВ (ИЗ НИХ 2 ОДНОГО РОДА) ИЗ РАЗНЫХ СЕМЕЙСТВ
VII. ЖИЗНЕННЫЙ ЦИКЛ ПЛАУНА БУЛАВОВИДНОГО И СЕЛАГИНЕЛЛЫ НАРИСУЙТЕ КОЛОСКИ И ЗАРОСТКИ ЭТИХ РАСТЕНИЙ
VIII. ХАРАКТЕРИСТИКА СЕМ. БОБОВЫЕ (МОТЫЛЬКОВЫЕ). УКАЖИТЕ ВАЖНЕЙШИЕ ДИКОРАСТУЩИЕ И КУЛЬТУРНЫЕ РАСТЕНИЯ ИЗ ЭТОГО СЕМЕЙСТВА (НЕ МЕНЕЕ 15 ВИДОВ)
IX. ПОНЯТИЕ О ЗОНАЛЬНОЙ И ИНТРАЗОНАЛЬНОЙ РАСТИТЕЛЬНОСТИ. ПРИВЕДИТЕ ПРИМЕРЫ
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
I. ПЛАСТИДЫ И ИХ ПИГМЕНТЫ, ФОТОСИНТЕЗ, НЕОБХОДИМЫЕ ДЛЯ НЕГО УСЛОВИЯ, ДЕЛЕНИЕ КЛЕТКИ
I.1 Пластиды и их пигменты
Пластиды это органеллы, характерные исключительно для растительных клеток. В них происходит первичный и вторичный синтез углеводов. Формы, размеры, строение и функции пластид различны. По окраске (наличию или отсутствию пигментов) Различают три типа пластид:
1. Зеленые хлоропласты – это органеллы фотосинтеза, относительно крупные структуры (5-10 мкм. в длину при диаметре 2-4 мкм.) овальной или дисковидной формы. Содержимое пластид называют стромой (матрикс). В строме содержатся белки, липиды, ДНК (кольцевая молекула), РНК, рибосомы и запасные вещества (липиды, крахмальные и белковые зерна). Кроме того, в строме содержатся также ферменты, участвующие в фиксации диоксида углерода. Наружная мембрана гладкая. Внутренняя – образует пластинчатые впячивания – тилакоиды, большая часть которых укладывается в виде стопки монет и образует граны. Хлоропласты образуются из пропластид – мельчайших сферических недеференцированных телец, котрые содержатся в растущих частях растения (в клетках зародыша, образовательной ткани) и имеют зеленый цвет, обусловленный присутствием фотосинтезирующим пигмента хлорофилла. Существует несколько видов хлорофилла.
Наиболее распространен хлорофилл а (найден у всех зеленых растений и цианобактерий) имеет голубовато-зеленый цвет
Хлорофилл б – желтовато-зеленый. Его молекула содержит на один атом кислорода больше и на два атома водорода меньше.
В процессе фотосинтеза хлорофиллу принадлежит ведущая роль. Он может поглощать солнечную энергию, запасать ее или передавать ее другим молекулам.
Кроме того в состав хлоропластов входят желтые пигменты – каротиноиды. Каротиноиды представляют собой высокомолекулярные углеводороды: оранжевый каротин С40Н56 и желтый ксантофилл С40Н56О2. Каротиноиды хлоропластов, а также синие, красные, бурые пигменты хроматофоров водорослей называют дополнительными, вспомогательными пигментами, поскольку энергия, поглощенная ими, может передаватться на хлорофилл а. Хлорофилл использует энергию красной части спектра, каротиноиды – синей. По своему строению каротиноиды являются либо углеводородами (каротины), либо окисленными углеводородами, т. е. Кислородсодержащими (каротинолы или ксантофиллы).Кроме того каротиноиды могут защищать молекулы хлорофилла от чрезмерного фотоокисления на ярком свету.
2. Лейкопласты – бесцветные округлые пластиды, в которых обычно накапливаются запасные питательные вещества, в основном крахмал. По строению лейкопласты мало отличаются от протопластид, из которых он образуется: двумембранная оболочка окружает бесструктурную строму. Внутренняя мембрана, врастая в строму, образует немногочисленные тилакоиды. В лейкопластах имеются ДНК, рибосомы, а также ферменты, осуществляющие синтез и гидролиз запасных веществ, в первую очередь крахмала. Лейкопласты, в которых синтезируется и накапливается запасной крахмал, называют амилопластами, белки – протеинопластами, масла – элайопластами. В одено лейкопласте могут накапливаться разные вещества. Запасной белок может откладываться форме кристаллов или аморфных включений, масла – в виде пластоглобул. Однако белки и масла в лейкопластах встречаются довольно редко. В амилопластах в связи с тилакоидами в строме возникают образовательные центры, вокруг которых в виде зерен откладывается вторичный запасной крахмал из растворимых углеводов, образовавшихся в хлоропластах в процессе фотосинтеза. Много амилопластов в клетках клубней картофеля, зерновок ржи, пшеницы и других органах растений где откладываются запасные вещества. В секреторных клетках они в комплексе с агранулярным ретикуломом участвую в синтезе эфирных масел.
3. Хромопласты – пластиды оранжево-красного и желтого цвета, образующиеся из лейкопластов и хлоропластов в результате накопления в их строме каротиноидов. Они встречаются в клетках лепестков (лютик, нарцисс, тюльпан, одуванчик), зрелых плодов (томат, тыква, арбуз, апельсин), редко – корнеплодов (морковь, кормовая свекла), а также в осенних листьях.
Хромопласты – конечный этап в развитии пластид. По форме накопления каротиноидов различают следующие типы хромопластов: глобулярный пигменты растворены в липидных пластоглобулах; фибриллярный – пигменты накапливаются в белковых нитях; кристалический – пигменты откладываются в виде кристалов.
Косвенное биологическое значение хромопластов в том, что ярко окрашенные плоды успешнее распространяются птицами и животными, а выделяющиеся яркой желто-красной окраской цветки привлекают насекомых – опылителей.
В филогенезе первичным исходным типом пластид являются хлоропласты, из которых в связи со специализацией органов произошли лейко- и хромопласты. В онтогенезе взаимопревращения пластид происходит иными путями. Наиболее часто хлоропласты превращаются в хромопласты при осеннем пожелтении листьев или созревании плодов. В природе этот процесс необратим. Лейкопласты могут превращаться в хлоропласты (позеленение верхней части корнеплода моркови, оказавшейся на поверхности почвы) или хромопласты. Хлоропласты могут при помещении растения в темноту превратиться в лейкопласты. Процесс этот обратим.
I.2 Фотосинтез, необходимые для него условия
Фотосинтез у зеленых растений – это процесс преобразования света в химическую энергию органических соединений, синтезируемых из диоксида углерода и воды. Процесс фотосинтеза представляет собой цепь окислительно-восстановительных реакций, совокупность которых подразделяют на две фазы – световую и темновую.
1. Световая фаза. Для этой фазы характерно то, что энергия солнечной радиации, поглощенная пигментами системы хлоропластов, преобразуется в электрохимическую.
При действии света на хлоропласт начинается электронный поток по системе переносчиков – сложных органических соединений, встроенных в мембраны тилакоидов. С переносом электронов по ЭТЦ сопряжено активное поступление протонов через тилакоидную мембрану из стромы внутрь тилакоида. В тилакоидном пространстве происходит увеличение концентрации протонов за счет расщепления молекул воды и в результате окисления электронного переносчика пластохинона на внутренней стороне мембраны. Когда протоны идут обратно по градиенту из тилакоидного пространства в строму, на наружной поверхности тилакоида с участием фермента АТФ-синтетазы из АДФ и фосфорной кислоты синтезируется АТФ, т. е. происходит фотосинтетическое фосфореилирование с запасанием энергии в АТФ, которая затем переходит в строму хлоропласта.
Заканчивается передача электронов следующим образом. Достигнув внешней поверхности мембраны тилакоида, пара электронов следует с ионом водорода, находящимся в строме. Оба электрона и ион водорода присоеденяются к молекуле переносчика водорода – НАДФ+ (никатиномидадениндинуклетидфосфат), который при этом переходит в свою востановленную форму
НАДФ•Н+Н+:
НАДФ++2Н++2е-→НАДФ•Н+Н+
Следовательно активированные световой энергией электроны используются на присоедининие атома водорода к его переносчику, т. е. на восстановление НАДФ+ в НАДФ•Н+Н+, который с наружной поверхности фотосинтетической мембраны переходит в строму.
В молекулах хлорофилла, утративших свои электроны, образовавшиеся электронные «дырки» действуют как сильный окислитель и отрывают электроны от молекул воды. Через ряд переносчиков эти электроны передаются на молекулу хлорофилла и заполняют «дырку». Внутри тилакоида происходит фотоокислние (фотолиз) воды, в результате которого выделяется свободный кислород, а также накапливаются ионы водорода
2Н2О→4Н++4е-+О2
Таким образом, во время световой фазы фотосинтеза происходят три процесса: образование кислорода вследствие разложения воды, синтез АТФ и образование атомов водорода в форме НАДФ•Н2. Кислород диффундирует в атмосферу, а АТФ и НАДФ•Н2 транспортируются в матрикс пластид и участвуют в процессе темновой фазы.
2.Темновая фаза фотосинтеза протекает в матриксе хлоропласта как на свету, так и в темноте и представляет собой ряд последовательных преобразований СО2, поступаещего из воздуха. Осуществляются реакции темновой фазы за счет энергии АТФ и НАДФ•Н2 и использовании имеющихся в пластидах пятиуглеродных сахаров, один из которых – рибулозодифосфат – является акцептором СО2. Ферменты связывают пятиуглеродный сахар с углекислым газом воздуха. При этом образуются соединения которые последовательно восстанавливаются до шестиуглеродной молекулы глюкозы.
Суммарная реакция фотосинтеза
6СО2+6Н2 энергия света С6Н12О6+6О2
![]() хлорофилл
хлорофилл
В процессе фотосинтеза кроме моносахаридов (глюкоза и др.), которые превращаются в крахмал и запасаются растением, синтезируются мономеры других органических соединений – аминокислоты, глицерин и жирные кислоты. Таким образом, благодоря фотосинтезу растительные, а точнее – хлорофиллсодержащие, клетки обеспечивают себя и все живое на Земле необходимыми органическими веществами и кислородом.
I.3 Деление клетки
Описано три способа деления эукариотических клеток: амитоз (прямое деление), митоз (непрямое деление) и мейоз (редукционное деление).
Амитоз – относительно редкий способ деления клетки. При амитозе интерфазное ядро делится путем перетяжки, равномерное распределение наследственного материала не обеспечивается. Нередко ядро делится без последующего разделения цитоплазмы и образуются двухъядерные клетки. Клетка, претерпевшая амитоз, в дальнейшем не способна вступать в нормальный митотический цикл. Поэтому амитоз встречается, как правило, в клетках и тканях, обреченных на гибель.
Митоз. Митоз, или непрямое деление, - основной способ деления эукариотических клеток. Митоз – это деление ядра, которое приводит к образованию двух дочерних ядер, в каждом из которых имеется точно такой же набор хромосом, что и был в родительском ядре.
В непрерывном процессе митотического деления различают четыре фазы: профазу, метафазу, анафазу и телофазу.
Профаза – самая длительная фаза митоза, когда происходит перестройка всей структуры ядра для деления. В профазе происходит укорочение и утолщение хромосом вследствие их спирализации. В это время хромосомы двойные (удвоение происходит в S-периоде интерфазы), состоят из двух хроматид, связанных между собой в области первичной перетяжки осбой структурой – цетромерой. Одновременно с утолщением хромосом исчезает ядрышко и фрагментируется (распадается на отдельные цистерны) ядерная оболочка. После распада ядерной оболочки хромосомы свободно и беспорядочно лежат в цитоплазме. Начинается формирование ахромативного веретена – веретена деления, которое представляет систему нитей, идущих от полюсов клетки. Нити веретена имеют диаметр около 25нм. Это пучки микротрубочек, состоящих из субъедениц белка тубулина. Микротрубочки начинают формироваться со стороны центриолей либо со стороны хромосом (в клетках растений).
Метафаза. В метафазе завершается образование веретена деления, которое состоит из микротрубочек двух типов: хромосомных, которые связываются с центромерами хромосом, и ценросомных (полюсных), которые тянутся от полюса к полюсу клетки. Каждая двойная хромосома прикрепляется к микротрубочкам веретена деления. Хромосомы как бы выталкиваются микротрубочками в область экватора клетки, т.е. располагаются на равном расстоянии от полюсов. Они лежат в одной плоскости и образуют так называемую экваториальную, или метафазную пластинку. В метафазе отчетливо видно двойное строение хромосом, соединенных только в области центромеры. Именно в этот период легко подсчитать число хромосом, изучать их морфологические особенности.
Анафаза начинается делением центромеры. Каждая из хроматид одной хромосомы становится самостоятельной хромосомой. Сокращение тянущих нитей ахроматинового веретена увлекает их к противоположным полюсам клетки. В результате у каждого из полюсов клетки оказывается столько же хромосом, сколько было их в материнской клетке, причем набор их одинаков.
Телофаза – последняя фаза митоза. Хромосомы деспирализуются, становятся плохо заметными. На каждом из полюсов вокруг хромосом воссоздается ядерная оболочка. Формируются ядрышки, веретено деления исчезает. В образовавшихся ядрах каждая хромосома состоит теперь всего из одной хроматиды, а не из двух.
Каждое из вновь образовавшихся ядер получило весь объем генетической информации, которым обладала ядерная ДНК материнской клетки. В результате митоза оба дочерних ядра имеют одинаковое количество ДНК и одинаковое число хромосом, такое же, как в материнском.
Цитокинез – после образования в телофазе двух новых ядер происходит деление клетки и формирование в экваториальной плоскости перегородки – клеточной пластинки.
В ранней телофазе между двумя дочерними ядрами, не достигая их, формируется цилиндрическая система волокон, называемая фрагмопластом, которая также как и волокна ахроматинового веретена, состоит из микротрубочек и связаны с ним. В центре фрагмопласта на экваторе между дочерними ядрами скапливаются пузырьки Гольджи, содержащие пектиновые вещества. Они сливаются друг с другом и дают начало клеточной пластинке, а их мембраны участвуют в построении плазмолемм по обеим сторонам пластинки. Клеточная пластинка закладывается в виде диска, взвешенного в фрагмопласте. Волокна фрагмопласта, видимо, контролируют направление движения пузырьков Гольджи. Клеточная пластинка растет центробежно по направлению к стенкам материнской клетки за счет включения в нее полисахаридов все новых и новых пузырьков Гольджи. Клеточная пластинка имеет полужидкую консистенцию, состоит из аморфного протопектина и пектатов магния и кальция. В это время из трубчатого ЭР образуются плазмодесмы. Расширяющийся фрагмопласт постепенно приобретает форму бочонка, позволяя клеточной пластинке расти латерально, пока она не соединится со стенками материнской клетки. Фрагмопласт исчезает, обособление двух дочерних клеток заканчивается. Каждый протопласт откладывает на клеточную пластинку свою первичную клеточную стенку.
Цитокинез с помощью клеточной пластинки происходит у всех высших растений и некоторых водорослей. У остальных организмов клетки делятся внедрением клеточной оболочки, которая постепенно углубляется и разделяет клетки.
Биологическое значение митоза состоит в строго одинаковом распределении между дочерними клетками материальных носителей наследственности – молекул ДНК, входящих в состав хромосом. Благодаря равномерному разделению реплицированных хромосом между дочерними клетками обеспечивается образование генетически равноценных клеток и сохраняется преемственность в ряду клеточных поколений. Это обеспечивает таки важные моменты жизнедеятельности, как эмбриональное развитие и рост организмов, восстановление органов и тканей после повреждения. Митотическое деление клеток является также цитологической основой бесполого размножения организмов.
Мейоз. Мейоз – это особый способ деления клеток, в результате которого происходит редукция (уменьшение) числа хромосом вдвое и переход клеток из диплоидного состояния (2n) в гаплоидное (n). Мейоз – единый, непрерывный процесс состоящий из двух последовательных делений, каждое из которых можно разделть на те же, что и в митозе, четыре фазы: профазу, метафазу, анафазу и телофазу. Обоим делениям предшествует одна интерфаза. В синтетическом периоде интерфазы до начала мейоза удваивается количество ДНК и каждая хромосома становится двухроматидной.
Первое мейотическое, или редукционное, деление.
Профаза I продолжается от нескольких часов до нескольких недель. Хромосомы спирализуются. Гомологичные хромосомы коньюгируют, образуя пары – биваленты. Бивалент состоит из четырех хроматид двух гомологичных хромосом. В бивалентах осуществляется кроссинговер – обмен гомологичными участками гомологичных хромосом, что приводит к их глубокому преобразованию. Во время коссинговера происходит обмен блоками генов, что объясняет генетическое разнообразие потомства. К концу профазы исчезает ядерная оболочка и ядрышко, формируется ахроматиновое веретено.
Метафаза I – биваленты собираются в экваториальной плоскости клетки. Ориентирование материнской и отцовской хромосомы из каждой гомологичной пары к одному или другому полюсу веретена деления является случайным. К центромере каждой из хромосом присоединяется тянущая нить ахроматинового веретена. Две сетринские хроматиды не разделяются.
Анафаза I – происходит сокращение тянущих нитей, и к полюсам расходятся двухроматидные хромосомы. Гомологичные хромосмы каждого из бивалентов уходят к противоположным полюсам. Расходятся случайно перераспределенные гомологичные хромосомы каждой пары (независимое распределение), и на каждом из полюсов собирается половинное число (гаплоидный набор) хромосом, образуется два гаплоидных набора хромосом.
Телофаза I – у полюсов веретена собирается одиночный, гаплоидный, набор хромосом, в котором каждый вид хромосом представлен уже не парой, а одной хромосомой, состоящей из двух хроматид. В короткой по продолжительности телофазе I восстанавливается ядерная оболочка, после чего материнская клетка делится на две дочернии.
Второе мейотическое деление следует сразу же после первого и сходно с обычным митозом (поэтому его часто называют митозом мейоза), только клетки, вступающие в него, несут гаплоидный набор хромосом.
Профаза II – непродолжительная.
Метафаза II – снова образуется веретено деления, хромосомы выстраиваются в экваториальной плоскости и центормерами прикрепляются к микротрубочкам веретена деления.
Анафаза II – осуществляется разделение их ценромер и каждая хроматида становится самостоятельной хромосомой. Отделившиеся друг от друга дочерние хромосомы направляются к полюсам веретена.
Телофаза II – завершается расхождение сестринских хромосом к полюсам и наступает деление клеток: из двух гаплоидных клеток образуются 4 клетки с гаплоидным набором хромосом.
Значение. Редукционное деление является как бы регулятором, препятствующим непрерывному увеличению числа хромосом при слиянии гамет. Не будь такого механизма, при половом размножении число хромосом удваивалось бы в каждом новом поколении. Т.е. благодаря мейозу поддерживается определенное и постоянное число хромосом во всех поколениях каждого вида растений, животных, протист и грибов. Другое значение заключается в обеспечении разнообразия генетического состава гамет как в результате кроссинговера, так и в результате различного сочетания отцовских и материнских хромосом при их расхождении в анафазе I мейоза. Это обеспечивает появление разнообразного и разнокачественного потомства при половом размножении организмов.
II. ВЫДЕЛИТЕЛЬНЫЕ СИСТЕМЫ РАСТЕНИЙ (железистые волоски, выделительные ходы, млечники и т.д.). ОХАРАКТЕРИЗУЙТЕ БАЛЬЗАМЫ, ЭФИРНЫЕ МАСЛА, СМОЛЫ, КАМЕДИ
II.1 Выделительные системы растений(железистые волоски, выделительные ходы, млечники и т.д.)
Выделительные системы в жизнедеятельности растений играют важную, разнообразную роль. В процессе их жизнедеятельности образуется ряд веществ с разнообразной химической природой, не участвующих в дальнейшем метаболизме.
Выделительные структуры растений характеризует, то что клетки выделительных тканей паренхимные, тонкостенные. Их ультраструктура связана с секретирующим веществом. В тканях, где синтезируются эфирные масла, смолы, каучук, имеется хорошо развитый агранулярный ЭР, слизи - аппарат Гольджи.
Выделительные ткани подразделяются на ткани внешней (наружной) и внутренней секреции.
Наружные выделительные структуры (внешняя секреция). Связаны эволюционно с покровными тканями. Они представлены следующими структурами: железистые волоски, желёзки, переваривающие желёзки, нектарники, осмофоры, гидатоды.
Железистые волоски и желёзки представляют собой трихомы эпидермы. Они состоят из живых клеток, обычно имеют удлиненную ножку из одной или нескольких клеток и одно- или многоклеточную головку. Клетки головки выделяют секрет под кутикулу. При разрыве кутикулы вещество изливается наружу, после чего может образовываться новая кутикула и накопиться новая капля секрета. Железистые волоски цветков герани, листьев и цветков душистого табака выделяют эфирные масла; сидячие головчатые волоски, образующие мучнистый налёт на листьях мари и лебеды, - воду и соли. Желёзки отличаются от волосков короткой ножкой из несекретирующих клеток и многоклеточной головкой. Они характерны у мяты, лаванды, полыни, черной смородины.
Нектарники обычно образуются на частях цветка, но могут встречаться и на других надземных органах растения. Они могут быть представлены отдельными поверхностными железистыми клетками или находиться в ямках (лютик), желобках, шпорцах (живокость), возвышаться в виде бугорков, подушечек (тыква, ива, яснотка белая). Выделительные клетки нектарников отличаются густой цитоплазмой и высокой активностью обмена веществ. К нектарнику может подходить проводящий пучок.
Осмофоры представляют собой или специализированные клетки эпидермы, или особые желёзки, где секретируются ароматические вещества. Выделение летучего секрета происходит в течение короткого времени и связано с использованием запасных веществ. Аромат цветка создается секрецией сложной смеси органических соединений, главным образом эфирных масел.
Гидатоды выделяют капельно-жидкую воду и растворенные в ней соли. При избытке воды и ослаблении транспирации через гидатоды происходит гуттация – выделение капель воды из внутренних частей листа на его поверхность. Специальной секреторной ткани здесь, как правило нет. Гидатоды могут иметь вид многоклеточных волосков, устьиц, потерявших способность регулировать величину своей щели, и, наконец представлять собой специальное образование из большого числа клеток, расположенных под водным устьицем.
Гидатоды в виде многоклеточных волосков имются у фасоли огненно-красной, а в виде водных устьиц встречается у манжетки, настурции. Наиболее сложно устроенные гидатоды находятся в зубцах листьев камнеломки, земляники, шиповника, чая. Здесь под водными устьицами образуется многоклеточная ткань эпитема, через которую вода передается от водопроводящих элементов к отверстиям устьиц.
Преваривющие желёзки на листьях насекомоядных растений, например росянки, венериной мухоловки и др., выделяют жидкость, содержащую пищеарительные ферменты и кислоты.
Внутренние выделительные структуры (внутренняя секреция). Вырабатывают и накапливают вещества, остающиеся внутри растения. Это могут быть отдельные секреторные клетки, рассеяные среди других тканей, как идиобласты. Они содержат различные вещества, особенно часто оксалат натрия в виде одиночных кристаллов, друз или рафид, бальзамы, танины, слизи и др.
Секреторные вместилища разнообразны по форме и происхождению. Схизогенные вместилища образуются вследствие расхождения клеток и формирования межклетника, выстланного живыми эпителиальными клетками и заполненного выделенными веществами. К ним относятся смоляные ходы хвойных растений. Лизигенные вместилища возникают в результате растворения группы клеток с продуктами секреции. Такие вместилища видны в кожуре плодов цитрусовых (апельсина, лимона, мандарина).
Млечники – особый тип выделительной ткани. Это живые клетки (нечленистые млечники) или ряды слившихся клеток (членистые млечники), пронизывающие всё растение. В зрелом млечнике протопласт занимает постенное положение, полость млечника занята млечным соком – латексом, млечные клетки неодревесневающие, эластичные. Латекс представляет собой эмульсию белого, реже оранжевого или красного цвета. Жидкая основа латекса – клеточный сок, в котором растворены или взвешены углеводы (крахмальные зерна у молочайных, сахар у астровых), белки (у фикуса), жиры танины, слизи, эфирные масла, каучук (более чем у 12 500 растений). Среди каучуконосов промышленное значение имеет тропическая гевея – Hevea (семейство Молочайные), в млечном соке которой содержится 40-50% каучука.
II.2 Характеристика бальзамов, эфирных масел, смол, камеди
Эфирные масла — летучие органические вещества самого разнообразного химического состава. Это бесцветные, иногда окрашенные, сильно летучие жидкости с весьма характерным, часто приятным ароматическим запахом и жгучим вкусом. Плохо растворяются в воде и хорошо в эфире, хлороформе, спирте и жирных маслах. Придают своеобразный запах различным частям растений, могут привлекать насекомых-опылителей, отпугивать травоядных. В мире известно 2500 эфиромасличных растений. Эфирные масла встречаются в различных частях растений — в цветках, листьях, плодах, а иногда и в подземных частях. Содержание их в растениях колеблется от следов до 20% (обычно 2—3%).
Смолы — твердые и полужидкие липкие органические вещества разнообразного химического состава. По химическому строению близки к эфирным маслам, часто содержатся в растениях вместе с ними. Они обладают характерным запахом, иногда сильно ароматическим. Некоторые смолы обладают противомикробным, дезинфицирующим и ранозаживляющим свойствами, многие являются балластными веществами. В медицинской практике применяются для приготовления пластырей, настоек, назначаются внутрь как слабительное средство.
Бальзамы - природные смолистые вещества, обогащенные эфирными маслами и др., которые образуются преимущественно в тропических и субтропических растениях.
Камеди (гумми), высокомолекулярные углеводы, составная часть соков и выпотов, выделяемых рядом растений при механических повреждениях и некоторых заболеваниях. Используется в промышленности как клей, стабилизатор суспензий и эмульсий и др., а также в виде слизей как обволакивающее средство.
III. ПОЧКИ, ТИПЫ ПОЧЕК ПО ПРОИСХОЖДЕНИЮ. ПРИВЕДИТЕ РИСУНОК ВЕГЕТАТИВНОЙ ПОЧКИВ ПРОДОЛЬНОМ РАЗРЕЗЕ
Почка (рис.1). Это зачаточный побег, междоузлия которого очень малы. Внутри почки заключен апекс побега – его меристематическая верхушка. Ниже него за счет выпячивания наружных слоев клеток экзогенно формируются листовые бугорки, превращающиеся в зачатки листьев – листовые примордии. Позже в их пазухах (обычно начиная с пазухи третьего-пятого от апекса примордия листа) образуются бугорки (зачатки почек следующего порядка), в связи с чем
Рисунок 1 их называют пазушными. Так как образование бугорков листьев чередуется с периодами, когда апекс растет в длину, восстанавливая свойственные ему величину и форму, листовые бугорки закладываются в определенном порядке и возникает характерное для вида листорасположение – филлотаксис. Период времени между заложением двух листовых бугорков называют пластохроном; период времени межу развертыванием двух соседних листьев – филлохроном.
Типы почек
В основу классификации почек положены различные признаки.
I. По наличию защитных чешуй:
1. Закрытые (защищенные)
2. Открытые (голые)
II. По составу и функции почки бывают:
1. вегетативные – состоят из зачаточного стебля, чешуек, зачаточных листьев и конуса нарастания;
2. генеративные – цветочные, состоят из зачаточного стебля, чешуй и зачатков цветка или соцветия (бузина красная);
3. вегетативно-генеративные (смешанные) – имеют зачатки листьев и цветков (яблоня, груша, вишня, слива и т.д.);
4. выводковые – метаморфизированные почки с зачатками придаточных корней, предназначены для вегетативного размножения. Они могут быть представлены луковичками (бульбиллами), развивающимися в пазухах листьев надземных побегов (некоторые лилии) или в соцветиях (некоторые луки); клубнелуковичками (гладиолусы); клубеньками в пазухах листьев (горец живородящий); почками, формирующимися в углублениях зубчиков листовой пластинки (каланхое).
III. По месторасположению на стебле:
1. Верхушечные.
2. Боковые.
1) Пазушные – развиваются в пазухах листьев и в свою очередь делятся на:
a) Одиночные;
b) сериальные почки, образующие вертикальный ряд (свойственны двудольным). Последовательность их заложения и развития:
· восходящая (жимолость, шалфей блестящий);
· нисходящая (грецкий орех, кирказон);
· бисериальная (двурядная)
c) коллатеральные почки, образующие горизонтальный ряд (характерны, г.о., для однодольных – гладиолус, крокус).
2) Придаточные (адвентивные) почки появляются на других органах: на листе (бриофиллюм, сердечник луговой, бегония), у корнеотпрысковых растений на корнях (вишня, слива, малина, хрен и т.д.).
IV. В КРУПНОМ МАСШТАБЕ НАРИСУЙТЕ СЕМЯЗАЧАТОКС ЗАРОДЫШЕВЫМ МЕШКОМ В ПРОДОЛЬНОМ РАЗРЕЗЕ. ИЗ КАКИХ ЧАСТЕЙ СЕМЯЗАЧАТКА РАЗВИВАЮТСЯТЕ ИЛИ ИНЫЕ ЧАСТИ СЕМЕНИ
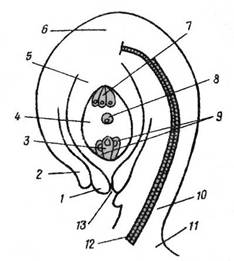
Рис. 2 Семязачаток
1, 2 – внутренний и наружный ингументы; 3 – яйцеклетка;
4 – зародышевый мешок; 5 – нуцеллус; 6 – халаза;
7 – антиподы; 8 – вторичное ядро (центральное); 9 – синергиды;
10 – фуникулюс; 11 – плацента;12 – проводящий пучок;
13 – пыльцевход (микропиле)
Из каких частей семязачатка развиваются те или иные части семени (рис. 2)
Зрелое семя состоит из зародыша, запасающих тканей и семенной кожуры, которое развивается обычно после оплодотворения из семязачатка.
Формирование зародыша происходит из зиготы (оплодотворенной яйцеклетки) – 3. Первое деление приводит к формированию двух клеток: базальной (со стороны макропиле) и терминальной (со стороны середины зародышевого мешка). Базальная клетка делится в поперечном направлении и формирует подвесок (суспензор), который прикрепляет зародыш к стенке зародышевого мешка и вдвигает его в полость зародышевого мешка. Самая верхняя крупная клетка-подвеска играет роль гаустория. Из терминальной клетки развивается собственно зародыш, в результате трёх её делений развивается шарообразное тело подзародыша, состоящее из мелких клеток. Далее зародыш на верхушке становится плоским, и по обе стороны закладываются два бугорка. У двудольных они развиваются симметрично, образуя две семядоли, у однодольных одна семядоля развивается энергичнее, а другая отстает в росте. Асимметрично развившаяся семядоля продолжает свой рост и занимает верхушечное положение, а другая остается рудиментарной.
Формирование эндосперма – оплодотворенное центральное ядро зародышевого мешка – 8, не проходя периода покоя делится и дает начало триплоидному эндосперму. Существует два типа эндосперма: нуклеарный и целлюлярный. При нуклеарном типе оплодотворенное централное ядро зародышевого мешка многократно делится, формируя множество триплоидных ядер. Клеточные стенки появляются поздно либо не формируются совсем. Нуклеарный тип эндосперма наблюдается у однодольных и некоторых двудольных. Целлюлярный тип формирования эндосперма отличается тем, что клеточные стенки образуются последовательно при каждом делении. Такой тип встречается у некоторых двудольных.
Формирование семенной кожуры(спермодерма) развивается из интегументов семязачатка. Она многослойна и присутствует в семени всегда. Протопласты наружних клеток кожуры отмирают, и клетки заполняются смолистым веществом, а стенки опробковевают, одревевсневают кутинизируются. Внутренние слои клеток остаются тонкостенными. Главные функции спермодермы – защита зародыша от механических повреждений, чрезмерного высыхания и преждевременного насыщения влагой и прорастания.
V. СПОСОБЫ РАСПРОСТРАНЕНИЯ ПЛОДОВ И СЕМЯН. ПРИВЕДИТЕ ПРИМЕРЫ. БИОЛОГОИЧЕСКАЯ РОЛЬ РАСПРОСТРАНЕНИЯ ПЛОДОВ И СЕМЯН
Различают следующие способы распространения плодов и семян:
· без участия посторонних агентов (семена и плоды крупных размеров);
· с помощью животных (сочные плоды, ягоду); ,
· с помощью ветра (плоды с крыльями и хохолками);
· с помощью воды (сухие плоды и семена);
· с помощью человека (все виды плодов и семян)
Автохория - распространение семян и плодов без участия посторонних агентов (семена и плоды крупных размеров)
Осуществляется в двух основных формах:
1) самопроизвольное опадение спелых диаспор (плодов, семян и т.п.) под собственной тяжестью (барохория);
2) активное разбрасывание семян (в радиусе от 1 до 15 м) при внезапном вскрывании плодов вследствии высокого и неравномерного напряжения в околоплоднике (автомеханохория). Эта форма автохории присуща и cочным плодам (некоторые тыквенные, бальзаминовые), но чаще сухим (бобовые, гераниевые, виды фиалки и др.).
Эпизоохория - распространение семян и плодов с помощью животных на их покровах - прежде всего на шерсти млекопитающих. Основные приспособления - наличие придатков, цепляющихся за шерсть:
Calamagrostis arundinacea - Вейник тростниковидный. Для этог