Эволюция выделительной системы беспозвоночных
Современный животный мир представляет собой результат длительной эволюции животных, в процессе которой по мере появления обособленной внутренней среды в живых организмах начинают обособляться и органы, предназначенные для поддержания постоянства её состава, и прежде всего – дыхательные и выделительные органы. Все клетки тела способны выделять жидкие или твёрдые продукты своего обмена. Этой тканевой экскреции вполне достаточно для тех животных, внутренняя среда которых мало обособлена от внешней, в силу чего продукты экскреции всех клеток поступаю непосредственно во внешнюю среду. С появлением обособленной внутренней среды появляются и органы, обладающие способностью поглощать из этой внутренней среды продукты обмена, поступающие туда из тканей, и выделять их наружу, т. е. специфические экскреторные органы. Первую ступень в развитии экскреторных органов представляют те случаи, когда часть клеток отнимает продукты экскреции от остальных клеток, без посредства общей для тех и других внутренней среды. И вероятно на этой ступени стоят более крупные кишечнополостные и большинство плоских червей.
В осмотическом отношении внутренняя среда типичных морских беспозвоночных не обособлена от окружающей морской воды. Но с переходом животного в воду неустойчивой солёности (литораль, эстуарии) или в пресную воду появляется необходимость в регуляции осмотического давления внутренней среды, и как приспособление к жизни в подобной среде возникают осморегулирующие органы. Последние первоначально совершенно независимы от экскреторных органов, но у вышестоящих животных обычно вступают с ним в тесную связь, образуя единый аппарат.
Экскреторные органы образуются за счёт всех пластов тела, но главным образом – за счёт фагоцитобласта. Наряду с ним большую роль в экскреции у всех животных играет периферический фагоцитобласт. Здесь экскреторную функцию берут на себя, с одной стороны – отдельные клетки и группы клеток неполяризованного фагоцитобласта, с другой стороны – целомический эпителий и стенки целомодуктов. Участие в экскреции кинобласта выражается больше всего в образовании за его счёт настоящих (эктодермальных) нефридиев. Более развитые типы нефридиев и экскреторные целомодукты, наряду с экскреторными придатками кишечника (мальпигиевы сосуды) образуют наиболее совершенные выделительные органы, с нефроцитами, расположенными в виде правильного эпителия и с выводным протоком.
Функция экскреции включает в себя предварительную переработку экскретов и выделение из организма нескольких различных категорий этих экскретов.
1. Зарождение выделительной системы и её дальнейшее развитие у беспозвоночных животных
1.1 Выделение продуктов метаболизма простейших
Простейшим, как и любым другим живым организмам, свойственны все функции: питание, обмен веществ, выделение, восприятие внешних раздражений и реакция на них, движение, рост, размножение и смерть.
Тело простейших устроено довольно сложно. В пределах одной клетки происходит дифференциация её отдельных частей, которые выполняют различные функции. Так, по аналогии с органами многоклеточных животных эти части простейших были названы органоидами или органеллами.
Выделительными органеллами простейших являются сократительные вакуоли. Кроме функции выделения накапливающихся в цитоплазме жидких продуктов метаболизма (обмена веществ), сократительная вакуоль освобождает простейшее от избытка воды, поступающей из окружающей среды в его тело. Т.е. уже у простейших функция экскреции объединяется с функцией осморегуляции, или поддержания постоянства внутренней среды организма. Морские простейшие, кроме инфузорий, и паразитические формы лишены сократительных вакуолей. Жидкие продукты обмена веществ выводятся у них диффузно всей поверхностью тела.
Строение сократительных вакуолей инфузорий:
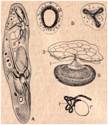
А – сократительные вакуоли и приводящие каналы Paramecium caudatum; Б - сократительные вакуоли инфузории Campenella umbellaria в состоянии диастолы и систолы; В - схема строения сократительной вакуоли Cycloposthium: 1 - вакуоль открывается наружу постоянным каналом, окружённым особыми мионемами-замыкателями; 2 - пелликула - плотный слой эктоплазмы. Г - сократительная вакуоль Paramecium trichium с извитым выводным каналом (1).
2. Выделение продуктов метаболизма многоклеточных
Возникновение многоклеточности было важнейшим этапом в эволюции всего царства животных. Размеры тела животных, ранее ограниченные одной клеткой, у многоклеточных значительно возрастают за счёт увеличения числа клеток. Тело многоклеточных состоит из нескольких слоёв клеток, не менее чем из двух. И среди клеток происходит разделение функций. Клетки дифференцируются на комплексы клеток, выполняющих одни и те же функции, образуют соответствующие ткани. А ткани, в свою очередь, образуют сложные органы и системы органов, обеспечивающие жизненные отправления животного.
Многоклеточность черезвычайно расширила возможности эволю-ционного развития животных и способствовала завоеванию ими всех возможных сред обитания.
Рассмотрим, как и за счёт чего происходит выделение экскрементов и продуктов метаболизма у представителей многоклеточных беспозвоночных животных:
2.1 Губки (Porifera)
Губки – наиболее низкоорганизованные асимметричные или радиально-симметричные многоклеточные животные, в основном обитающие в морях и океанах. Они находятся на клеточном уровне организации. Это означает, что их тело представляет собой скопление рыхло расположенных, в разной степени дифференцированных клеток, выполняющих определённые функции. Однако эти клетки ещё не образуют настоящих тканей и органов.
Выделение у губок, как и дыхание, осуществляется диффузно. Жидкие продукты метаболизма и экскременты из пищеварительных вакуолей хоаноцитов – цилиндрических или шаровидных клеток, образующих жгутиковые камеры, и амёбоцитов – крупных блуждающих клеток, участвующих в переваривании пищи и осуществляющих фагоцитоз, поступют через систему каналов в атриальную полость, а оттуда с током воды выбрасываются через оскулум наружу.
2.2 Кишечнополостные (Coelenterata)
Тип кишечнополостных относится к низшим многоклеточным животным, которые, однако, по целому ряду признаков состоит выше губок. Это приемущественно морские животные, и лишь немногие из них живут в пресных водах.
Выделение экскрементов у кишечнополостных происходит через ротовое отверстие. Также выделительную функцию приписывают и кишечным порам кишечнополостных, но с физиологической стороны это предположение недостаточно обосновано. Зато морфологически они являются вероятными гомологами целомодуктов, которые у многих животных несут выделительную функцию.
2.3 Плоские черви (Plathelminthes)
К типу плоских червей относятся наиболее низкоорганизованные трёхслойные билатеральные животные, ведущие разнообразный образ жизни. У плоских червей имеется ряд особенностей в их строении и биологии, развившихся под влиянием условий жизни. Такой особенностью плоских червей является появление у них новой системы органов, отсутствующей у кищечнополостных – выделительной системы.
Органы выделения впервые возникают у турбуллярий (ресничных червей). И у различных турбуллярий выделительные органы развиты неодинаково. Они представлены системой сильно разветвлённых каналов, часто образующих перемычки, или анастомозы. Наиболее тонкие канальцы замыкаются слепо концевыми, или терминальными, клетками, а главные каналы открываются выделительными отверстиями. Терминальные клетки грушевидной формы, часто со звёздчатыми отростками, расположены они непосредственно в паренхиме. Внутри клеток имеется полость, в которой помещается пучок длинных ресничек (1). Пучок ресничек находится в непрерывном колебательном движении, напоминающем колебания пламени свечи, за что эти клетки называют пламенными клетками. Полость терминальной клетки продолжается в её отростке. Это и есть начало выделительного канала. Далее к отростку клетки примыкает ряд удлинённых клеток, сквозь которые проходит канал. Канальцы, отходящие от близко расположенных пламенных клеток, соединяются в более крупные протоки, затем эти протоки впадают в ещё более крупные, открывающиеся одним или несколькими отверстиями наружу.
Описанные органы выделяют из тела избыток воды, а также жидкие продукты диссимиляции. Продукты распада органических веществ диффузно проникают из паренхимы в полость выделительной клетки и движением мерцательного пламени гонятся по каналам, которые также выстланы ресничками, и, наконец, выделяются наружу.
Важнейшей чертой выделительных органов турбуллярий, как и всех плоских червей, является наличие особых терминальных клеток, замыкающих выделительные каналы. Такого типа выделительные органы беспозвоночных называют протонефридиями.
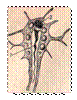
Выделительная система плоских червей - протонефридиального типа:

Выделительная система турбуллярий:
А - расположение главных выделительных каналов в теле прямокишечной турбуллярии Gyratryx hermaphrodites; Б - схема строения протонефридиев:
1 – главные продольные выделительные каналы, 2 – разветвления канальцев, 3 – звёздчатые клетки, 4 – мерцательное пламя, 5 – выделительные отверстия
![]()
Выделительная система трематод
1 – главные боковые каналы выделительной системы, 2 – протонефридиальные канальцы, заканчиваю-щиеся мерцательными клетками,3 – мочевой пузырь
![]()
Выделительная система ленточных червей:
1 – продольные боковые каналы выделительной системы,
2 – разветвления выделительных каналов, заканчивающиеся звёздчатыми клетками,
3 – поперечные перемычки между продольными каналами
2.4 Немертины(Nemertini)
Водные, почти исключительно морские свободноживущие, реже паразитические животные. У немертин впервые появляется хорошо развитая кровеносная система. Накапливающиеся в крови конечные продукты обмена веществ поступают в выделительную систему, устроенную по типу протонефридиев. Это пара трубок, лежащих по бокам тела, чаще лишь в передней его половине, и открывающихся наружу выводными отверстиями. Каждый ствол даёт от себя много побочных канальцев, оканчивающихся клетками с мерцательным пламенем. Своими концами канальцы глубоко вдаются в стенку боковых кровеносных сосудов, облегчая этим диффузию конечных продуктов обмена веществ из крови в выделительную систему.
![]()
Немертины: 1 – выделительные органы
2.5 Круглые (Первичнополостные) черви (Nemathelminthes)
Этот тип характеризуется либо отсутствием выделительной системы, либо она представлена видоизменёнными кожными железами, либо она протонефридиального типа.
Выделительная система первичнополостных на примере класса собственно круглых червей (Nematoda):
Один из самых многочисленных классов животного мира, свободноживущих и паразитирующих. Выделительная система нематод своеобразна. Она состоит из одноклеточных кожных (гиподермальных) желез, заменивших протонефридии, которые исчезли в связи с утратой нематодами ресничных образований, и выполняющих также функцию осморегуляции. Выделительные органы расположены в гиподерме и образованы обычно одной или двумя, реже многими клетками и эти органы называются шейными железами. У части свободноживущих нематод она массивна и имеет короткий выделительный проток (её дополняют более мелкие железы, лежащие по бокам тела) и кроме шейных желез, у них имеются терминальные, или хвостовые, выделительные кожные железы. У большинства почвенных и паразитических нематод шейная железа связана с длинными экскреторными каналами, расположенными в боковых утолщениях гиподермы. Например, у лошадиной аскариды (Parascaris equorum) в боковых валиках гиподермы проходят два выделительных канала, замкнутых на задних концах. Эти каналы, проходят к переднему концу тела, загибаются на брюшную сторону, сливаются вместе в короткий непарный канал и открываются на брюшной стороне выделительным отверстием. Весь этот орган представляет собой одну гигантскую клетку: выделительные каналы проходят в её отростках, а ядро расположено в цитоплазме левого канала.
Через боковые каналы выводятся вырабатываемые в теле жидкие продукты выделения. Но у нематод имеются также особые фагоцитарные органы, в которых задерживаются и накапливаются различные нерастворимые продукты обмена веществ и посторонние тела, проникающие в организм, например бактерии. Следовательно, нерастворимые частицы, накопившиеся в фагоцитарных органах, не удаляются из тела, а только устраняются таким образом из жизненного круговорота организма.
Такие органы называются « почками накопления»; они имеют вид крупных звёздчатых клеток (у лошадиной аскариды их четыре). Они расположены в полости тела по ходу боковых выделительных каналов, в передней трети тела. Кроме того, в них обнаружен гемоглобин. Предполагают, что фагоцитарные органы служат центрами, регулирующими потребление свободного кислорода нематодами.

Одноклеточные выделительные органы паразитических нематод:
А – Porrocaecum; Б – Contracaecum; В – Parascaris equorum;
1 – ядро выдели-тельной клетки; 2 – выделительное отверстие; Г – вскрытый передний конец Parascaris equorum: 1 – кишка, 2 – фагоцитарные органы, 3 – боковые выделительные каналы.
2.6 Кольчатые черви (Annelida)
Тип кольчатых червей, или кольчецов, представляет собой очень важную группу для понимания эволюции высших беспозвоночных животных.
Основным признаком внешнего строения кольчецов является метамерия, или сегментация тела. Метамерия кольчецов выражается не только во внешней, но и во внутренней организации, в повторяемости многих внутренних органов. Это касается и выделительной системы: функция выделения выполняется сегментарными органами – нефридиями (метанефридиями). Обычно в каждом сегменте имеется одна пара нефридиев.
Метанифридиями называются открытые выделительные органы, в отличие от замкнутых протонефридиев. Метанефридии расположены метамерно и попарно, но так, что каждый нефридий начинается в одном (переднем) сегменте, а заканчивается выделительным отверстием в следующем (заднем).
Выделительная функция метанефридиев двоякого рода. Стенка нефридиального канала, особенно средней части, пронизана сетью кровеносных капилляров. Из крови в нефридиальный канал поступают жидкие продукты диссимиляции. С другой стороны, через ресничный аппарпт воронки вместе с частью целомической жидкости выносятся экскреты, которые предварительно накапливаются в амёбоидных клетках целома, а также в особых клетках, обладающих фагоцитарной функцией и расположенных в определённых местах целомического эпителия. По мере накопления зёрнышек твёрдых экскретов эти клетки вместе с содержащимися в них экскретами выносятся наружу. Таким образом, развитие метанефридиев тесно связано с участием целома в выделительных процессах.
Выделительная система класса малощетинковых кольчецов (Polychaeta):
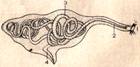
Метанефридий:
1 – воронка; 2 – начало нефридиального канала; 3 – нефридиальный канал с ресничками; 4 – конечная часть нефридиального канала, открывающегося порой.
У большинства высокоорганизованных полихет органами выделения служат метанефридии.
Метанефридий начинается более или менее расширенной воронкой – нефростомом, усаженной ресничками и открывающейся в полость сегмента. От нефростома начинается нефридиальный канал, который проходит через диссепимент в следующий сегмент. Здесь нефридиальный канал образует более или менее сложный клубок, подходит к стенке тела и открывается и открывается выделительным отверстием наружу. К метанефридиям подходят кровеносные сосуды. В просвете нефридиального канала обычно имеются реснички.
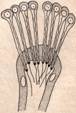
Конец одной из ветвей протонефридия
На первый взгляд метанифридии совершенно не похожи на протонефридии. Однако эти два типа выделительных органов генетически между собой связаны, и метанефридии развились из протонефридиев. Протонефридии есть у полихет. Личинка полихет – трохофора – имеет настоящие протонефридии с выделительными каналами, замкнутыми терминальными клетками. У многих полихет имеются протонефридии несколько особого типа. У них нефридиальный канал в терминальной части замкнут, и кончается группой клеток, напоминающих булавки с расширенными головками. В расширенной части этих клеток помещаются ядро и цитоплазма, а в трубчатой ножке проходит канал, в котором находится колеблющийся жгут, доходящий до начала нефридиального канала. Такие нефридиальные клетки получили название соленоцитов. Функция соленоцита та же, что и у звёздчатой клетки протонефридиев плоских червей, т.е. осморегуляция. Далее, у некоторых полихет имеются открытые нефридии, но ещё без воронки, а с пучком мерцающих длинных ресничек, Это как бы следующий этап в эволюции выделительных органов. Очевидно, при превращении протонефридиев в метанефридии сначала произошла редукция замкнутой терминальной части и образование открытого канала, а затем развитие воронки.
У полихет, содержащих в сегментах гонады, нефридии соединяются с половыми протоками. Исходной формой половых протоков у многощетинковых были, по-видимому, половые воронки, или целомодукты, - короткие каналы мезодермальной природы, открывающиеся одним концом наружу, другим (воронкой) в целом. У некоторых полихет произошло прирастание половых воронок к главному каналу имевшегося в каждом сегменте протонефридия; получились смешанные нефридии, или нефромиксии, играющие одновременн роль и выделительных органов, и выводных протоков для половых продуктов.
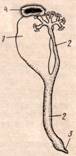
Нефромиксии полихет: 1,4 - половая воронка, 2 – нефридиальный канал, 3 – наружное отверстие.
Кроме нефридиев есть и другие образования, играющие выделительную роль. В некоторых местах, главным образом на стенках кровеносных сосудов, перитонеальный эпителий состоит из крупных клеток, содержащих множество жёлтых зёрен, - это накопляющиеся в клетках нерастворимые продукты обмена веществ (гуанин или соли мочевой кислоты). Наполнившись экскретами, эти как их называют, хлорагогенные клетки отмирают, а содержимое их поступает в целом и оттуда через нефридии – наружу.
Выделительная система малощетинковых кольчецов (Oligochaeta) также представлена типичными метанефридиями и хлорагогенными клетками.
Выделительная система класса пиявок (Hirudinea):
Пиявки – сильно изменённые потомки Oligochaeta. Это свободноживущие хищники или гораздо чаще эктопаразиты, нападающие на других нередко крупных животных и питающиеся их кровью.
Выделительная система пиявок – метанефридиального характера, но сильно изменена. Выделительные каналы расположены метамерно, но обыкновенно отсутствуют в части передних и задних сегментов. Так у медицинской пиявки, у которой число сегментов равно 33, нефридиев всего 17 пар. Нефридий представляет извитой канал, составленный на большем своём протяжении из одного ряда клеток, пронизанных внутриклеточным просветом. Канал открывается на боковой стенке тела выводным отверстием. Главная особенность нефридиев пиявок состоит в том, что канал их на внутреннем конце замкнут слепо. В тесном соседстве со слепым концом канала находится особая мерцательная воронка, смотрящая своим широким концом в один из остатков целома, чаще всего в узкую поперечную лакуну. Суженным концом воронка вдаётся в небольшой слепой мешочек – резервуар, примыкающий, к слепому концу нефридиального канала. Судя по всему, у пиявок произошло разобщение воронки с остальной частью метанефридия. Воронкой захватываются амёбоциты, нагруженные продуктами обмена, и переводятся в резервуар, а через его стенки экскреты, по-видимому, осмотически просачиваются в канал нефридия. Стенки части целомических каналов (лакун) покрыты скоплениями зернистых клеток, совокупность которых называется «ботриоидной тканью». Эти скопления сходны с хлорагогенными клетками олигохет.
Строение медицинской пиявки (Hirudo medicinalis): 1 – канал метанефридия, 2 – мочевой пузырь, 3 – боковой лакунарный канал
![]()
Выделительная система типа кольчатых червей:

Поперечный разрез многощетинкового червя:
1 - канал нефридия, 2 – воронка нефридия

Поперечный разрез тела дождевого червя ( Lumbricus terrestris): 1 – метанефридий
2.7 Моллюски (Mollusca)
Моллюски представляют собой самый многочисленный тип животных после членистоногих. В подавляющем большинстве это типичные водные животные и только сравнительно немногие приспособились к жизни на суше.
Выделительные органы моллюсков метанефридиального типа. Почки, или нефридии, это настоящие целомодукты. Каждый из них открывается воронкой, или нефростомом, в перикардиальную полость. Другой конец нефридия открывается нефропорой в мантийную полость, т.е. во внешнюю среду. Проходящие в тесном соседстве с почками приносящие жаберные сосуды (с венозной кровью) образуют многочисленные бахромчатые слепые выпячивания (венозные придатки), которыми вдаются в стенки почек, этим облегчается извлечение почками продуктов обмена веществ из крови.
Выделительная система типа моллюски:

Выделительная система неопилины (Neopilina) из класса моноплакофоры: 1 – почки, 2 – наружные отверстия почек, 3 – анальное отверстие.

Выделительная система класса хитоны (Loricata):
1 - почки, 2 – наружные отверстия почек, 3 – воронка, открывающаяся в перикар-диальную полость, 4 – анальное отверстие.

Выделительная система брюхоногих (Gastropoda): 1 - анальное отверстие, 2 – почка.

Выделительная система каракатицы (Sepia) из класса головоногих (Cephalopoda): 1 – почки, 2 – наружное отверстие почек.
2.8 Иглокожие (Echinodermata)
Иглокожие - обширная группа морских донных животных, большей частью свободноподвижных, реже прикреплённых ко дну.
Специальные органы выделения у иглокожих отсутствуют. Значительная часть образующихся в теле продуктов обмена веществ выводится наружу при помощи амёбоидных клеток, рассеянных в жидкости, наполняющей все полости тела. При впрыскивании в целом мелко растёртой туши амёбоциты нагружаются зёрнами краски и выходят из тела через его покровы. При этом они избирают наиболее тонкие участки стенки тела, а именно кожные жабры; через них выходят це6лые скопления амёбоидных клеток, нагруженных тушью и нормальными продуктами выделения. Часть экскретов прямо откладывается в коже и других тканях в виде скоплений жёлтых зёрен и т.п. Запас амёбоидных клеток в организме необходимо постоянно пополнять. Для этой цели служат особые лимфатические органы: тидемановы железы и осевой орган. Тидемановы железы имеют вид небольших телец, расположенных парами на околоротовом амбулякральном кольцевом канале по бокам полиевых пузырей и каменистого канала. Осевой орган прилегает к каменистому каналу в виде продолговатого мешка, состоящего из губчатой ткани (видоизменённая кровеносная лакуна): в ячеях губчатой ткани лежат многочисленные округлые клетки, активно делящиеся и производящие таким способом новые амёбоциты.
У класса голотурии (Holothuroidea) (у безногих) имеется ряд интересных приспособлений выделительного характера. Вдоль кишечного мезентерия перитонеальный эпителий образует ряд нескольких десятков довольно крупных мерцательных вороночек, слепо замкнуты у своего основания. При иньекции туши в полость тела можно наблюдать, как вороночки заполняются множеством полостных амёбоцитов, нагрузившихся зёрнами туши. Амёбоциты не выводятся через воронки, но склеиваются слизистым выделением их в так называемые «бурые тела», которые выталкиваются из вороночек в полость тела. Таким образом воронки служат главным образом как бы для концентрации накопляющихся в теле экскретов, напоминая в этом отношении «урночки» класса сипункулид типа кольчатых червей.
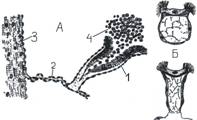
Урночки сипункулид и голотурий:
А - разрез через урночку голотурии Chiridota pelliucida:
1 –урночка, 2 – мезентерий кишечника, 3 – стенка тела, 4 – фагоциты, наполненные тушью; Б – прикреплённая и плавающая урночки сипункулид.
2.9 Членистоногие (Arthropoda)
Членистоногие составляют большинство животного населения земного шара как по числу видов, так и по абсолютной численности. Среди беспозвоночных членистоногие представляют собой наиболее высокоорганизованных и чрезвычайно разнообразных животных (не считая класса головоногих моллюсков).
Многообразие и процветание членистоногих объясняется тем, что в процессе эволюции предки членистоногих под влиянием условий жизни приобрели ряд особенностей, развитие которых обеспечивало чрезвычайно широкие возможности расселения этой группы животных и приспособления их к самым разнообразным условиям существования. Представители членистоногих живут не только в морях, океанах и пресных водах, но и почти повсюду на суше. Это единственный тип беспозвоночных, большинство которого являются наземными, сухопутными животными.
Выделительная система членистоногих представлена видоизменёнными целомодуктами – коксальными железами или же особыми органами, возникшими в пределах типа членистоногих, а именно мальпигиевыми сосудами.
Выделительная система класса ракообразные (Crustacea):
Ракообразные составляют существенную часть водной фауны. Встречаются как планктонные, так и бентосные формы.
Выделительная система раков почти утратила метамерный характер. Имеются две пары выделительных железистых органов (антеннальные, или зелёные, железы) – видоизменённых целомодуктов. Строение обеих пар приблизительно одинаково. Каждый орган состоит из концевого мешочка и отходящего от него извитого канала с железистыми стенками; канал делает несколько петлеобразных изгибов и затем открывается наружу, иногда образуя перед этим заметное расширение – мочевой пузырёк. Одна пара открывается у основания антенн (антеннальные железы), другая – у основания второй пары нижних челюстей (максиллярные железы). Установлено, что концевой слепо замкнутый мешочек представляет собой сохранившийся от эмбрионального состояния участок целома, а отверстие, сообщающее пузырёк с каналом, - воронку целомодукта, только лишённую мерцательных ресничек. Лишь в очень редких случаях (отряд Leptostraca) обе пары выделительных желез присутствуют одновременно. Обычно же одна из них функционирует на личиночных стадиях и потом редукцируется, заменяясь во взрослом состоянии другой. При этом все раки,кроме подкласса Malacostraca, во взрослом состоянии имеют максиллярные железы, а Malacostraca – антеннальные (зелёные) железы.
Выделительная система класса многоножки (Myriapoda):
Этот класс состоит исключительно наземных, иногда довольно крупных членистоногих.
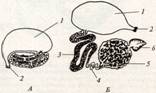
Антеннальная железа речного рака:
А – в естественном состоянии; Б - в расправленном виде:
1 – мочевой пузырёк, 2 – выделительная пора, 3 – белый канал, 4 прозрачный канал, 5 – зелёный канал, 6 – целомический мешочек.
![]()
Коксальные железы сенокосца Garella variegate:
1 – основания ходных ног, 2 – трахейный ствол, 3 - выделительное отверстие, 4 – перерезанный выводной канал железы, 5 – стигма, 6 – резервуар железы (мочевой пузырь).
На границе между средней и задней кишками в кишечник впадают 1 или 2 пары длинных слепозамкнутых на свободном конце трубок – мальпигиевых сосудов (1). В эпителии сосудов и их просвете скапливаются конкреции мочевой кислоты; последняя у многоножек, так же как и у насекомых представляет собой главный продукт выделения. Кроме мальпигиевых сосудов выделительную роль играют и другие образования, прежде всего лимфатические железы в виде неправильных клеточных тяжей, расположенных либо вдоль мальпигиевых сосудов, либо вдоль брюшного кровеносного сосуда, или вдоль брюшной нервной цепочки. Они служат для захвата и накопления твёрдых продуктов выделения и фагоцитируют впрыснутые в полость тела твёрдые частицы. Кроме того, участие в выделении принимает жировое тело. Полость тела – миксоцель – многоножек во многих местах заполнена неправильными тяжами и скоплениями клеток. Совокупность таких скоплений и называется жировым телом. В его клетках наблюдаются многочисленные капли жира, а также конкреций мочевой кислоты. Жировое тело служит не только для накопления питательного материала, но и для выделения.
Выделительная система класса паукообразные (Arachnida):
Паукообразные – высшие хелицеровые членистоногие. Они дышат посредством лёгких или трахей и обладают помимо выделительного аппарата в виде ветвящихся мальпигиевых сосудов энтодермического происхождения, лежащих в брюшке (в просветах которых накапливается гуанин – основной продукт выделения паукообразных) - коксальными железами – парными мешковидными образованиями мезодермальной природы, лежащими в двух (реже в одном) сегментах головогруди. Коксальные железы хорошо развиты у зародышей и в молодом возрасте, но у взрослой особи более или менее атрофируются. Вполне сформированные коксальные железы состоят из концевого эпителиального мешочка, петлевидно извитого канала и более прямого выводного протока с мочевым пузырьком и наружным отверстием. Концевой мешочек отвечает ресничной воронке целомодукта, отверстие которой замкнуто остатком целомического эпителия. Коксальные железы открываются у основания 3-й или 5-й пары конечностей.

Схема организации паука:
1 – мальпигиевы сосуды, 2 – основания ходных ног.
Выделительная система класса насекомые (Insecta):
Насекомые представляют собой самый большой класс животных в современной фауне. Они широко расселились поп материкам и приспособились к самым различным условиям существования. Насекомые – наиболее высокоорганизованные членистоногие, но среди них разные группы достигли различной степени сложности организации.
Функции органов выделения у насекомых выполняются рядом образований. Но основными органами выделения насекомых являются мальпигиевы сосуды, отходящие от пищевого канала на границе между средней и задней кишками и имеющие эктодермальное происхождение. Мальпигиевы сосуды иногда ветвятся, но чаще представляют собой длинные неветвящиеся трубочки, лежащие вдоль кишечника в миксоцеле. Они слепо замкнуты на одном конце и другим концом открываются в кишечник на границе средней и задней кишок. Их полость выстлана низким однослойным эпителием. Количество сосудов колеблется от 2 до 200 и более или менее постоянно у представителей разных систематических групп. Жидкие продукты выделения в виде мочевой кислоты, уратов и других соединений попадают из мальпигиевых сосудов в задний отдел кишечника и выделяются наружу. Иногда мальпигиевы сосуды обладают некоторой подвижностью, что обеспечивает постоянную смену омывающей их гемолимфы.

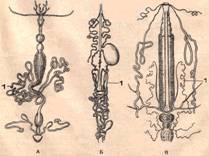
Мальпигиевы сосуды различных насекомых:
А – жужелицы; Б – бабочки; В – гусеницы бабочки; Г – чёрного таракана: 1 – мальпигиевы сосуды
Кроме мальпигиевых сосудов, выделительную функцию у насекомых выполняет также жировое тело. Этот орган занимает промежутки между органами и бывает особенно развит на определённых стадиях развития, например у гусениц бабочек старших возрастов. Это связано с тем, что в клетках жирового тела накапливаются питательные вещества в виде капелек жира, которые затем используются в период метаморфоза. Во многих жировых клетках и в особых клетках жирового тела – нефроцитах – обнаруживаются отложения мочекислых солей. Поглощая продукты диссимиляции, клетки жирового тела никуда их не отдают, они функционируют в данном случае как «почки накопления».
Помимо этого, выделительную функцию несут и некоторые другие органы. Таковыми, по-видимому, являются группы перикардиальных клеток, лежащих по бокам сердца. Возможно, такую же роль играют находимые в жировом теле и в полости тела особые клетки – эноциты. У низших насекомых (ногохвосток, тизанур и двухвосток) имеются нижнегубные выделительные железы, по-видимому, гомологичные метанефридиям полихет.
метаболизм беспозвоночный выделительный экскреторный
Заключение
Выделительная система беспозвоночных животных разных уровней организации имеет относительное сходное строение, и в процессе эволюции идут по пути усложнения. Выделительными органеллами простейших являются сократительные вакуоли. Губки иглокожие и кишечнополостные не имеют специальных выделительных органов, и продукты обмена веществ переходят у них из внутриклеточной жидкости во внешнюю среду путём диффузии за счёт клеток хоаноцитов и амёбоцитов. Наиболее примитивные животные - это плоские черви и немертины обладают специализированными выделительными органами, у них имеются протонефридии, заканчивающиеся звёздчатыми (пламенными) клетками. У представителей кольчатых червей в каждом сегменте тела имеется по паре специализированных выделительных органов – метанефридиев, произошедших от протонефридиев. Моллюскам также характерны выделительные органы метанефридиального типа - почки, или нефридии, являющиеся настоящими целомодуктами. Среди беспозвоночных членистоногие представляют собой наиболее высокоорганизованных животных, для них характерно появления видоизменённых целомодуктов – коксальных, антеннальных желез или же особых органов, возникших в пределах типа членистоногих, а именно мальпигиевых сосудов. Нужно сказать о том, что в пределах разных типов и классов беспозвоночных животных, помимо основных характерных органов выделения, присутствуют и другие дополнительные образования, также несущие функцию выделения продуктов метаболизма.
Библиографический список:
1. Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. Изд. 2-е. Гос. изд. « Советская наука» - М.; 1952
2. Билич Г.Л., Крыжановский В.А. Биология полный курс. 3т. Зоология 4-е издание / М.: «Оникс», 2004
3. Гинецианская Т.А. Трематоды их жизненный цикл, биология и эволюция. М.: Наука, 1968
4. Догель В.А. Зоология беспозвоночных: Учебник для ун-тов / Под ред. проф. Полянского Ю.И.- 7-е изд., перераб. и доп. – М.: Высш. школа, 1981
5. Мантейфель Б.П. АНССР Институт эволюционной морфологии и экологии животных им. Северцова _ М.: Наука, 1984
6. Натали В.Ф. Зоология беспозвоночных. Учебник для студентов биол. фак. пед. ин-тов/ Под ред. Проф. Сазоновой О.Н. - 3-е изд., перераб. и доп. – М.: «Просвещение», 1975
7. Шапкин В.А., Тюмасева З.И. , Машкова И.В., Гуськова Е.В. Практикум по зоологии беспозвоночных: Учебное пособие для студ. высш. пед. учеб. заведений / - М.: Издательский центр «Академия», 2003