Инфракрасная спектроскопия и спектроскопия кругового дихроизма. Методы определения вторичной структуры белков
1. Спектры кругового дихроизма белков
1.1 Явление кругового дихроизма
1.2 Методы анализа спектров кругового дихроизма белков
1.3 Работа с пакетом программ STRUCTURE по анализу спектров КД белков
2. Инфракрасные спектры поглощения белков
2.1 Поглощение белков в ИК-области
2.2 Методы анализа ИК-спектров белков
2.3 Работа с пакетом программ STRUC по анализу ИК-спектров белков
Список литературы
Введение
Хромофоры белковых молекул (то есть химические группы в молекуле белка, ответственные за поглощение света на определенных длинах волн) можно разделить на три класса: пептидные группы, боковые группы аминокислотных остатков и простетические группы. Спектроскопические методы исследования вторичной структуры белка основаны на изучении спектров именно пептидных хромофоров, поскольку конформация пептидных групп и определяет тот или иной тип вторичной структуры белка - a-спираль, b-структуру и др. Изучение поглощения света пептидными группами белка обычно проводится в ультрафиолетовом и в инфракрасном диапазонах. Как показывают эксперименты, простая адсорбционная спектроскопия белков в неполяризованном ультрафиолетовом свете мало пригодна для анализа вторичной структуры белка. Более ценную информацию можно извлечь из спектров кругового дихроизма белка. Инфракрасные спектры поглощения белка также пригодны для анализа его вторичной структуры (1). Ниже будет рассмотрено применение методов измерения кругового дихроизма и инфракрасной спектроскопии для анализа вторичной структуры белка.
1. Спектры кругового дихроизма белков1.1 Явление кругового дихроизма
Белки, как практически все биологические молекулы, вследствие своей пространственной асимметрии обладают оптической активностью. При прохождении через оптически активную среду плоскополяризованный свет становится эллиптически поляризованным. Эллиптичность света q является одной из мер оптической активности. Она определяется как арктангенс отношения малой и большой осей эллипса. Другим параметром, характеризующим оптическую активность, является отклонение большой оси эллипса от направления поляризации падающего света, называемое оптическим вращением (или дисперсией оптического вращения) j.
Если представить плоскополяризованную волну Е в виде суммы двух волн противоположной круговой поляризации Е=ЕL+ЕR, то можно показать, что величина j пропорциональна разности показателей преломления среды для этих волн nL-nR, а величина q - разности коэффициентов экстинции eL-eR. Таким образом, оптическое вращение и появление эллиптической поляризации у плоскополяризованного света при прохождении его через оптически активную среду можно объяснить различным замедлением (nL¹nR) и поглощением (eL¹eR) двух его составляющих ЕL и ЕR, поляризованных по кругу. Разность Dn=nL-nR называют круговым двулучепреломлением, а разность De=eL-eR - круговым дихроизмом. Зависимости этих величин от длины волны называют спектрами дисперсии оптического вращения (ДОВ) и кругового дихроизма (КД).
На самом деле, ДОВ и КД являются проявлениями одного и того же физического явления, а их спектры можно выводить один из другого. Поэтому на практике достаточно измерять лишь один из этих двух спектров. Спектры КД более удобны для использования на практике, поскольку содержат узкие, хорошо разрешимые полосы. Этим объясняется то, что в настоящее время метод измерения КД используется гораздо более широко, чем ДОВ, несмотря на то, что он требует гораздо более сложного экспериментального оборудования.
КД легко измерить путем попеременного пропускания через образец лево - и правополяризованного по кругу света и регистрации соответствующей разницы поглощений, поскольку эллиптичность выходящего из оптически активного образца света обычно очень мала, и ее точное измерение весьма затруднительно. Однако, разность поглощений обычно пересчитывают в значения эллиптичности. Для того, чтобы можно было сравнивать результаты, полученные при исследовании разных образцов, пользуются значениями так называемой молярной эллиптичности:
(q) = 100q / Cl= 3300 De, (1.1.1)
где С - молярная концентрация, а l - длина оптического пути.
В случае белков главной целью измерения спектров КД является определение содержания в них вторичных структур разных типов. Если доля ароматических аминокислот в белке не очень велика, его оптическая активность в области от 180 до 240 нм определяется главным образом полипептидным остовом. Многочисленные эксперименты показали, что алифатические боковые группы аминокислотных остатков белка также не дают заметного вклада в спектр КД в этой области. Следовательно, в первом приближении белковую молекулу можно рассматривать просто как комбинацию участков полипептидным остова, находящихся в конформациях a-спирали, b-структуры и беспорядочного клубка.
Поглощение света пептидной группой
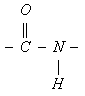
в ультрафиолетовом диапазоне определяется электронными переходами в ее электронных оболочках. В этом процессе основное участие принимают три молекулярных орбитали пептидной группы: n-орбиталь - несвязывающая орбиталь, на которой располагается неподеленная пара 2py-электронов атома кислорода, p-орбиталь - связывающая орбиталь, на которой располагаются 2pz-электрон атома кислорода и 2pz-электрон атома углерода, в значительной степени делокализованные по атомам кислорода, углерода и азота, и p*-орбиталь - разрыхляющая орбиталь, на которой в основном состоянии электроны отсутствуют. Два электронных перехода с наименьшей энергией наблюдаются при возбуждении электрона с n-орбитали на p*-орбиталь (n®p* переход) и с p-орбитали на p*-орбиталь (p®p* переход). n®p* переходу в пептидах соответствует слабая полоса поглощения 210-220 нм, а p®p* переходу гораздо более сильная полоса с максимумом вблизи 190 нм (характерная для a-спиральной конформации).
КД различных типов вторичной структуры белка можно оценить по результатам измерения КД гомополипептидов известной конформации (например, поли-L-лизина), после чего определить вклад каждой из структур в спектр КД исследуемого белка. Однако, такой подход имеет ряд больших недостатков. Во-первых, участки упорядоченной вторичной структуры модельных гомополипептидов имеют значительно большую длину, чем длина типичных участков в глобулярных белках. Во-вторых, их конформация может сильно отличаться от конформации, наблюдаемой у элементов вторичной структуры реальных белков. Кроме этого, среди гомополипептидов нельзя найти "стандартов" для b-изгибов. И, наконец, хотя вклад в КД от взаимодействий между хромофорами уменьшается как квадрат расстояния между ними, должен существовать определенный вклад от взаимодействия между участками с различной вторичной структурой. Эти взаимодействия нельзя адекватно смоделировать, рассматривая протяженные гомополимеры. Поэтому на практике спектры КД гомополипептидов не используются. Вместо этого в качестве базисных берут спектры КД белков, структура которых известна из данных рентгеноструктурного анализа. Различные подходы к анализу исследуемого спектра КД на основе этого базисного набора определяют различия между методами, которые будут описаны ниже.
1.2 Методы анализа спектров кругового дихроизма белковМетод "эталонных спектров" (2,3). Методы предсказания вторичной структуры белков по их спектрам КД основаны на предположении о том, что спектры КД различных структурных форм, составляющих белковую молекулу, дают аддитивный вклад в спектр КД белка в целом. Это можно записать в следующем виде:
![]() (1.2.1)
(1.2.1)
где ![]() - спектр КД белка (зависимость эллиптичности от длины волны света),
- спектр КД белка (зависимость эллиптичности от длины волны света), ![]() - идеализированный "эталонный спектр" - спектр КД, соответствующий i-ой структурной форме, участвующей в образовании вторичной структуры белка,
- идеализированный "эталонный спектр" - спектр КД, соответствующий i-ой структурной форме, участвующей в образовании вторичной структуры белка, ![]() - доля этой формы во вторичной структуре, причем
- доля этой формы во вторичной структуре, причем
![]() и
и![]() . (1.2.2)
. (1.2.2)
Эталонные спектры ![]() для всех структурных форм могут быть вычислены на основании набора базисных спектров КД (спектров белков с известной вторичной структурой - коэффициентами
для всех структурных форм могут быть вычислены на основании набора базисных спектров КД (спектров белков с известной вторичной структурой - коэффициентами ![]() ) с помощью метода наименьших квадратов и формулы (1.2.1), примененной к каждому базисному спектру. После этого экспериментальный спектр КД исследуемого белка с помощью того же метода наименьших квадратов может быть аппроксимирован по формуле (1.2.1) с использованием вычисленных эталонных спектров. При этом вклад каждого из эталонных спектров будет равен доле соответствующей ему структурной форме в общей структуре белка Такой подход к анализу спектров КД белков был впервые использован в работе (2). Ниже будет более подробно рассмотрена модификация этого метода (3).
) с помощью метода наименьших квадратов и формулы (1.2.1), примененной к каждому базисному спектру. После этого экспериментальный спектр КД исследуемого белка с помощью того же метода наименьших квадратов может быть аппроксимирован по формуле (1.2.1) с использованием вычисленных эталонных спектров. При этом вклад каждого из эталонных спектров будет равен доле соответствующей ему структурной форме в общей структуре белка Такой подход к анализу спектров КД белков был впервые использован в работе (2). Ниже будет более подробно рассмотрена модификация этого метода (3).
Принимая в рассмотрение в качестве структурных классов a-спираль (H), b-структуру (b), b-изгиб (t) и “неупорядоченную” форму (R), можем написать:
![]() . (1.2.3)
. (1.2.3)
Суммируя экспериментальные данные, вместо ![]() в уравнение (1.2.3) вводят величину
в уравнение (1.2.3) вводят величину ![]() , учитывающую зависимость эталонного спектра, соответствующего a-спирали, от числа аминокислотных остатков, образующих ее:
, учитывающую зависимость эталонного спектра, соответствующего a-спирали, от числа аминокислотных остатков, образующих ее:
![]() , (1.2.4)
, (1.2.4)
где ![]() и
и ![]() - эталонные спектры для a-спирали из аминокислотных остатков и для a-спирали “бесконечной” длины, а k - так называемый фактор длины цепи (
- эталонные спектры для a-спирали из аминокислотных остатков и для a-спирали “бесконечной” длины, а k - так называемый фактор длины цепи (![]() ). Согласно теоретическим расчетам оптической активности a-спирали и экспериментальным данным, спектр КД a-спирали
). Согласно теоретическим расчетам оптической активности a-спирали и экспериментальным данным, спектр КД a-спирали ![]() в диапазоне 185-240 нм может быть разложен на три независимых оптически активных составляющих (n®p*, p®p||*, p®p^*), которые можно описать гауссовскими зависимостями:
в диапазоне 185-240 нм может быть разложен на три независимых оптически активных составляющих (n®p*, p®p||*, p®p^*), которые можно описать гауссовскими зависимостями:
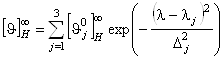 , (1.2.5)
, (1.2.5)
где ![]() и
и ![]() - положение максимума и полуширина j-ой гауссовской линии в спектре КД a-спирали, а
- положение максимума и полуширина j-ой гауссовской линии в спектре КД a-спирали, а ![]() - максимальное значение эллиптичности “бесконечной” a-спирали на длине волны
- максимальное значение эллиптичности “бесконечной” a-спирали на длине волны ![]() . В окончательном виде для спектра КД белка можно написать следующее выражение:
. В окончательном виде для спектра КД белка можно написать следующее выражение:
![]() , (1.2.6)
, (1.2.6)
где
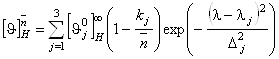 . (1.2.7)
. (1.2.7)
Здесь ![]() - среднее число аминокислот на a-спиральный участок цепи в молекуле белка.
- среднее число аминокислот на a-спиральный участок цепи в молекуле белка.
Параметры ![]() ,
, ![]() ,
, ![]() и
и ![]() в уравнении (1.2.7) были найдены на основе спектра КД миоглобина. Они имеют следующие значения:
в уравнении (1.2.7) были найдены на основе спектра КД миоглобина. Они имеют следующие значения:
j |
|
|
|
|
| 1 | 223.4 | 10.8 | -3.73×10 | 2.50 |
| 2 | 206.6 | 8.9 | -3.72×10 | 3.50 |
| 3 | 193.5 | 8.4 | +10.1×10 | 2.50 |
Подобные работы: