Менделевская генетика
На протяжении долгой истории научной (в большей степени натурфилософской) мысли преобладающими были метафизические представления о наследственности и изменчивости. Уже в далекие времена, когда начался период одомашнивания различных животных, предпринимались попытки улучшить их полезные качества. Решая эти задачи, человечество интуитивно опиралось на биологические закономерности наследования. Начиная с трудов Гиппократа, Аристотеля, Платона, других древнегреческих врачей и философов, появляются первые теоретические объяснения явлению наследственности. В XVIII и XIX столетиях изучением проблемы наследования занимались такие выдающиеся ботаники и врачи, как И.Кельейтер, Т. Найт, Ш.Ноден, П. Мопертюи и другие. Было показано, что признаки родителей, в том числе и нежелательные, например болезни, передаются через половые клетки; описано преобладание у гибрида одного признака над другим. Однако основоположником науки генетики, открывшим главные законы наследования признаков, является гениальный чешский ученый Г. Мендель.
Главная заслуга Менделя состоит в разработке и использовании гибридологического метода для анализа явлений наследования.
До открытий Менделя признавалась теория так называемой слитной наследственности. Суть этой теории состояла в том, что при оплодотворении мужское и женское «начало» перемешивались, «как краски в стакане воды», давая начало новому организму. Мендель заложил фундамент представлений о дискретном характере наследственного вещества и о его распределении при образовании половых клеток у гибридов.
Основные результаты семилетних экспериментов по изучению законов наследования Мендель опубликовал в бюллетене общества естествоиспытателей в г. Брюнне (ныне г. Брно, Чехия) в 1866 г. Исследование называлось «Опыты над растительными гибридами». Однако эта публикация не привлекла внимания современников. Только через 35 лет, в 1900 г., когда законы наследования были вновь открыты сразу тремя ботаниками — К. Корренсом, Э. Чермаком и другими) они получили всеобщее признание. К настоящему времени правильность законов Менделя подтвержден на громадном числе растительных и животных организмов, в том числе и на человеке. Открытие Менделем законов, отражают процесс передачи наследственной информации и принципа дискретности (генной детерминации наследственных признаков), явилось первым экспериментальным доказательством существования наследственности как реального материального явления.
Гибридологический метод Г. Менделя
Гибридологический метод — это система специальных скрещиваний для получения гибридов с целью анализа характера наследования признаков.
Мендель объектом своих экспериментов выбрал растение, полностью отвечающее поставленной задаче: оно имело надежную защиту от посторонней пыльцы во время цветения и обладало нормальной плодовитостью. Такими растениями были различные сорта самоопыляющегося посевного гороха (Pisum sativum).
Суть разработанного Менделем метода состоит из нескольких основных постулатов.
1. Подбор исходных «константно различающихся" родительских пар.
Для скрещивания использовались растения, отличавшиеся некоторыми признаками: например, окраской цветка (у одного растения пурпурная, у другого — белая), длиной стебля (у одного растения около 2 м, у другого — до 60 см) и т.д. В своих экспериментах Мендель изучал наследование 7 альтернативных пар признаков: окраски цветка, расположения цветков (пазушное или концевое), высоты растений, характера поверхности горошин (гладкая или морщинистая), окраски горошин (желтая или зеленая) и т.д. В каждом поколении Мендель вел учет альтернативных признаков отдельно по каждой паре. До начала экспериментальных скрещиваний Мендель в течение нескольких лет проводил работу на получение «чистых линий», т.е. сортов, постоянно и устойчиво воспроизводящих анализируемый признак. (Термин «чистые линии» возник много позднее, датский генетик — селекционер В.Иогансен так назвал группу особей с однородной наследственностью.)
2. Количественный анализ полученных гибридов, отличающихся по отдельным признакам от каждой родительской пары.
3. Индивидуальный анализ потомства от каждого скрещивания в ряду поколений.
Революционное новшество данного методического приема заключалось в учете и анализе потомства, полученного путем размножения всех без исключения гибридных особей.
4. Применение статистических методов оценивания результатов эксперимента.
Г.Мендель имел в практику генетического анализа систему записей скрещивания, в которой символ P обозначает родителей (лат. parenta - родители); F — потомков от скрещивания (лат. filii — дети). Позднее стали использовать нижний цифровой индекс при символе F для обозначения последующих поколений. Например, F![]() - обозначает потомство от скрещивании родительских форм; F
- обозначает потомство от скрещивании родительских форм; F![]() — обозначает потомство от скрещивания гибридов первого поколения и т.д.; символ «x» означает скрещивание особей.
— обозначает потомство от скрещивания гибридов первого поколения и т.д.; символ «x» означает скрещивание особей.
Моногибридное скрещивание
В первом поколении, полученном от скрещивания родительских форм, имеющих различия только по одной паре признаков (например, гладкие и морщинистые горошины; высокие и низкие стебли; окрашенные и белые цветы), были получены гибриды, у которых проявился признак только одного из родителей (только гладкие горошины, только высокие растения, только окрашенные цветы и т.д.). Никаких переходных (смешанных) форм растений по другим признакам не было обнаружено. Признак одного из родителей, проявляющийся у гибрида, Мендель назвал доминантным (от лат. dominare — господствовать, властвовать), а парный, не проявившийся признак был назван рецессивным (от лат. гесеssus — уступающий, отступающий назад) признаком. В последующем явление преобладания у гибридов первого поколения признака одного из родителей стали называть законом единообразия гибридов первого поколения, или первым законом Менделя.
Анализ потомков гибридов первого поколения, полученных путем самоопыления, позволил обнаружить, что наряду с доминантными формами вновь появляются растения с признаками, отсутствовавшими в поколении гибридов первого поколения, причем в строгих числовых отношениях. Таким образом, родительские признаки у гибридов первого поколения не исчезали и не смешивались. Мендель предположил, что эти признаки присутствовали у гибридов первого поколения в скрытом виде, но не проявлялись, почему он и назвал их рецессивными признаками. Оказалось, что по всем парам изученных признаков особи с доминантными и рецессивными признаками появлялись в соотношении 3:1. Появление доминантных и рецессивных форм в потомстве, полученном от самоопыления гибридов первого поколения, и составляет сущность чакона расщепления, или второго закона Менделя.
На основании полученных результатов Мендель пришел к следующим выводам:
1. Так как исходные родительские сорта не давали расщепления, у гибрида первого поколения (с доминантным признаком) должно быть два наследственных задатка (в современной терминологии — два аллеля).
2. Гибриды первого поколения содержат по одному задатку, полученному от каждого из родительских растений через половые клетки.
3. Наследственные задатки у гибридов первого поколения не сливаются, а сохраняют свою индивидуальность.
Для подтверждения своих выводов Мендель предпринял анализирующее, или возвратное, скрещивание — скрещивание гибрида первого поколения с рецессивной родительской особью. В потомстве от этого вида скрещивания он, как и ожидал, получил как доминантные, так и рецессивные формы в соотношении 1:1. Это подтвердило, что отдельные наследственные задатки при образовании половых клеток попадают в различные гаметы. Таким образом, гибрид первого поколения образует два типа половых клеток: клетки, содержащие наследственный задаток, определяющий доминантный признак, и клетки, содержащие наследственный задаток, определяющий рецессивный признак. В этом смысле каждая половая клетка «чистая», т.е. содержит один, и только один, аллель из пары (правило чистоты гамет). Распределение контрастных наследственных задатков в соотношении 1:1 является всеобщим биологическим законом, лежащим в основе всех других закономерностей наследования признаков.
В настоящее время особь, которая имеет два различающихся аллеля в каждом локусе гомологичных хромосом и которая, следовательно, образует два типа половых клеток, называется гетерозиготой (от греч. heteros — другой, различный + zygotos — соединение, пара). Особь, в каждой гомологичной хромосоме которой находятся идентичные аллели и которая, следовательно, образует только один тип половых клеток, называется гомозиготой (от греч. homos — тот же самый + zygotos — соединение, пара). Используя буквенную символику, введенную Г.Менделем для обозначения каждого наследственного задатка (для доминантного — прописная буква «А», для рецессивного — «а» строчная), можно изобразить схему опытов.
На рис. V.1 показано, что все потомки от данного вида скрещивания унаследовали признак только одного из родителей, т. е. они единообразны. Поскольку каждый из родителей образовал только один тип гамет (только «А» или только «а»), все потомки оказались гетерозиготами.
На рис.V.2 видно, что, судя по внешнему виду, на каждые три растения с доминантным признаком приходится одна особь с рецессивным признаком, в то время как по наследственным потенциям наблюдается иное расщепление — 1АА : 2Аа : 1аа.
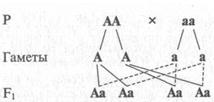
Рис. V.I. Схема, иллюстрирующая закон единообразия гибридов первого поколения. Все потомство — гетерозиготы
Действительно, при дальнейшем размножении потомков от скрещивания гибридов первого поколения (Аа) Мендель наблюдал, что рецессивные особи (аа) и одна треть доминантных (АА) не дают расщепления признаков в потомстве. Однако 2/з доминантных потомков (Аа) вновь образуют при самоопылении доминантные и рецессивные формы в соотношении 3:1.
Существенным результатом скрещивания двух гетерозигот явилось появление внешне сходных растений (с доминантными признаками, например, с гладкими горошинами; окрашенными цветками и т.д.), в то же время различавшихся по наследственным задаткам (АА и Аа). Оказалось, что организмы с одинаковыми признаками могут иметь различную генетическую конституцию.
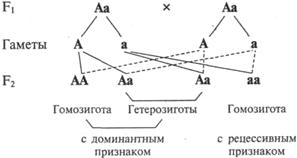
Рис. V.2. Схема, иллюстрирующая закон расщепления.
Таким образом, стали различать расщепление по внешнему проявлению признаков, которое соответствует отношению 3:1 (расщепление по фенотипу), и расщепление по наследственным задаткам, выражаемое отношением 1:2:1 (расщепление по генотипу). Термины «генотип» и «фенотип» были предложены В.Иогансеном в 1909 г. Генотип — это наследственная конституция особи как совокупность генов организма. Термин «генотип» часто используется для обозначения генетической конституции особи применительно к анализу наследования конкретного признака. Фенотип (от греч. phaino — являю + typos — образец) — совокупность признаков организма (анатомических, физиологических, биохимических, психических и т.д.). Термин «фенотип» часто используется для обозначения конкретного признака (внешнего или внутреннего) как результата проявления конкретного гена.
На основании полученных результатов можно заключить, что при моногибридном скрещивании двух гетерозиготных особей расщепление в потомстве по фенотипу соответствует двум классам признаков и может быть выражено отношением 3:1; расщепление по генотипу соответствует отношениям 1:2:1, т.е. возникает три генотипических класса.
Наследственные задатки, определяющие парные альтернативные признаки (обозначенные Менделем латинскими буквами А, а), впоследствии стали называть аллельной парой, а отдельный конкретный фактор — аллелью. В настоящее время аллель определяют как одно из альтернативных состояний одного и того же гена.
Типы межаллельных взаимодействий
Рассмотренные в предыдущем разделе явления доминантности и рецессивности представляют собой один из примеров взаимодействия различных аллелей. Однако вскоре после вторичного открытия законов Менделя были обнаружены факты, указывающие на существование и других видов межеаллельных отношений.
Так, в некоторых случаях при моногибридном скрещивании расщепление по фенотипу и генотипу могут совпадать. Это имеет место в ситуации, когда у гетерозигот отмечается промежуточное выражение признака, т.е. гибрид F, не воспроизводит полностью ни один из родительских признаков. Например, при скрещивании гомозиготной доминантной формы растения — ночной красавицы (Mirabilis jalapa), имеющей красные цветки (АА), и гомозиготной рецессивной формы, имеющей белые цветки (аа), в первом поколении гибридов наблюдаются только розовые цветы (генотип Аа), отличающиеся от обеих родительских форм. Во втором поколении (F![]() ) отмечается расщепление по окраске цветка на три фенотипи-ческих класса соответственно расщеплению генотипов: 1АА (красные) : 2Аа (розовые): 1аа (белые).
) отмечается расщепление по окраске цветка на три фенотипи-ческих класса соответственно расщеплению генотипов: 1АА (красные) : 2Аа (розовые): 1аа (белые).
Тот случай, когда признак у гетерозигот имеет промежуточное между доминантным и рецессивным проявление, получил название неполного доминирования. Явление неполного доминирования широко распространено в природе, в том числе и у человека. Оно может касаться как нормальных, так и патологических признаков. Например, у человека по типу неполного доминирования наследуется одна из форм анофтальмии (отсутствие глазных яблок). У доминантных гомозигот (АА) глазные яблоки нормальных размеров; у гетерозигот (Аа) глазные яблоки уменьшены в размере, но зрение сохранено, а у рецессивных гомозигот (аа) глазные яблоки отсутствуют. В браке двух индивидов с уменьшенными размерами глазных яблок в среднем 1/4 часть детей будет иметь нормальные глаза (АА), 1/2 часть — уменьшенные глазные яблоки (Аа) и 1/4 часть детей рождается без глазных яблок (аа).
Иная ситуация возникает в случае, когда отношения доминантности и рецессивности отсутствуют и оба аллеля проявляются в фенотипе. Подобное взаимодействие аллелей было названо совместным доминированием, или кодоминированием.
Явление кодоминирования можно проиллюстрировать на примере наследования групп крови системы MN у человека. Известно, что группы крови системы MN находятся под контролем одного гена (L), имеющего два аллеля (L![]() и L
и L![]() ). Если один из родителей имеет группу крови ММ (гомозигота по аллелю М), а другой — NN, то в эритроцитах их детей (гетерозиготы MN) выявляются как антигены М, так и антигены N. Подобные гены носят название кодоминантных генов.
). Если один из родителей имеет группу крови ММ (гомозигота по аллелю М), а другой — NN, то в эритроцитах их детей (гетерозиготы MN) выявляются как антигены М, так и антигены N. Подобные гены носят название кодоминантных генов.
Определяя термин «аллель» как альтернативное состояние одного гена, ученые вслед за Менделем остановились на рассмотрении только двух его состояний. На самом деле один и тот же ген может иметь более чем два состояния. Например, ген А может видоизменяться (мутировать) в несколько состояний: а![]() , а
, а![]() , а
, а![]() , и т.д. до многих десятков. Подобные аллели, т.е. изменения одного и того же гена, называются серией множественных аллелей. У диплоидного организма она может быть представлена только одной парой — двумя любыми аллелями. Например, АА, Аа
, и т.д. до многих десятков. Подобные аллели, т.е. изменения одного и того же гена, называются серией множественных аллелей. У диплоидного организма она может быть представлена только одной парой — двумя любыми аллелями. Например, АА, Аа![]() , а
, а![]() а
а![]() , а
, а![]() , а
, а![]() и т.д. Наследование членов серии множественных аллелей подчиняется рассмотренным законам. Каждый из членов серии множественных аллелей может полностью доминировать один над другим, например в порядке убывания: А > а
и т.д. Наследование членов серии множественных аллелей подчиняется рассмотренным законам. Каждый из членов серии множественных аллелей может полностью доминировать один над другим, например в порядке убывания: А > а![]() > а
> а![]() > а
> а![]() и т. д. В этой ситуации каждый последующий член аллельной серии будет доминировать над всеми остальными, кроме предыдущих.
и т. д. В этой ситуации каждый последующий член аллельной серии будет доминировать над всеми остальными, кроме предыдущих.
Наследование групп крови системы ав0
Группа крови системы АВО (читается как «а, б, ноль») контролируется одним аутосомным геном, т.е. геном, расположенным в одной из аутосомных (не половых) хромосом. Локус этого гена обозначается латинской буквой I (от слова «изогемагглютиноген»), а его три аллеля 1![]() , 1
, 1![]() и 1
и 1![]() обозначаются для краткости, как А, В и 0. Аллели А и В — кодоминантны по отношению друг к другу, и оба доминантны по отношению к аллелю 0. При сочетании различных аллелей могут образоваться 4 группы крови, различающихся между собой иммунологическими свойствами как эритро-
обозначаются для краткости, как А, В и 0. Аллели А и В — кодоминантны по отношению друг к другу, и оба доминантны по отношению к аллелю 0. При сочетании различных аллелей могут образоваться 4 группы крови, различающихся между собой иммунологическими свойствами как эритро-
| Генотип | Фенотип | |
| Группа крови | Антитела сыворотки | |
I | 0(I) |
|
I | A(II) |
|
I | A(II) |
|
I | B(III) |
|
I | B(III) |
|
I | AB(IV) | 0 |
Связь между генотипом и фенотипом групп крови системы АВ0
цитов, так и сыворотки (табл. V.1). Эритроциты содержат антигены (агтлютиногены), а в сыворотке находится вещество агглютинин (от лат. agglutinatio — склеивание), называемое антителом.
Определение групповой принадлежности человека по системе АВ0 осуществляется при проведении реакции агглютинации (рис. V.3).
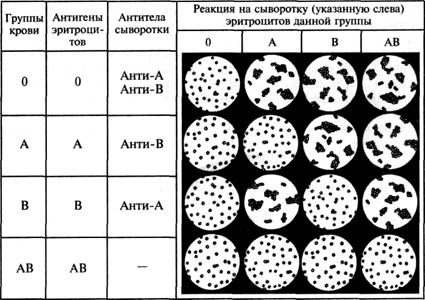
Рис. V.3. Взаимодействие эритроцитов индивидов с группами крови 0, А, В и АВ с антителами сыворотки (анти-А и анти-В).
Знать групповую принадлежность крови человека — необходимое условие безопасного проведения переливания крови. Термин «универсальный донор» обозначает лицо с 0(I) группой крови, так как его эритроциты не могут быть агглютинированы сывороткой ни одного реципиента. «Универсальный реципиент» — индивид с AB(IV) группой крови, сыворотка которого не может агглютинировать эритроциты какого-либо донора.
Полигибридные скрещивания
Основные закономерности, открытые Г.Менделем, касались наследования и расщепления только по одной паре альтернативных признаков (при моногибридном скрещивании). На следующем этапе Менделя интересовал вопрос, какими признаками будет обладать потомство от скрещивания родительских форм, различающихся одновременно несколькими признаками.
Гибриды, полученные от скрещивания особей, различающихся одновременно по двум парам альтернативных признаков, носят название дигибридов. Рассмотрим результаты классического опыта Менделя по дигибридному скрещиванию.
Для скрещивания были отобраны растения, которые имели гладкие желтые горошины (оба признака доминантные), и растения с морщинистыми зелеными горошинами (оба признака рецессивные). Согласно первому закону все потомство было единообразно: дигибридные растения давали только гладкие желтые семена. В потомстве от самоопыления 15 дигибридных растений было получено 556 горошин: из них 315 гладких желтых, 108 гладких зеленых, 101 морщинистых желтых и 32 горошины были морщинистые зеленые. Это соотношение близко к соотношению 9:3:3:1 и отражает относительные частоты 4 классов фенотипов.
При анализе по каждому признаку в отдельности (только по форме или только по окраске) расщепление гибридов в F![]() соответствовало соотношению 3:1.
соответствовало соотношению 3:1.
Проследим расщепление по признаку формы семян. Гладких горошин было 315 + 108, что составило в сумме 423 гладких. Морщинистых горошин было 101 + 32, что составило 133 семян. Отношение 423 к 133 было близко к уже известному отношению 3:1. То же самое отношение было получено при анализе расщепления по признаку окраски. Желтых горошин вне зависимости от их формы было 416 (315 + 101), а количество зеленых составило 140 горошин (108 + 32). Отношение также близко к 3:1.
Полученные результаты свидетельствовали, что расщепление по каждой паре аллелей при дигибридном скрещивании происходит как два независимых события. Таким образом, соотношение фенотипов при дигибридном скрещивании представляет собой результат случайного или независимого объединения результатов двух моногибридных скрещиваний. Этот вывод отражает сущность третьего закона Менделя — закона независимого комбинирования генов.
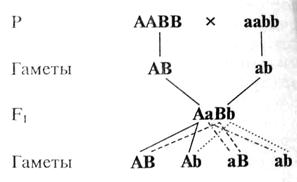
Обозначив доминантный аллель, определяющий желтую окраску, буквой А и рецессивный аллель, определяющий зеленую окраску, буквой а, буквой В — доминантный аллель, контролирующий развитие гладкой формы горошины, и соответственно рецессивный аллель, контролирующий развитие морщинистой формы, буквой b, дигибридное скрещивание можно записать следующим образом (рис. V.4).
Для того чтобы представить фенотипические и генотипические классы потомков дигибридов первого поколения, воспользуемся так называемой решеткой Пеннета, которая позволяет установить все возможные сочетания мужских и женских гамет (табл. V.2).
Легко убедиться, что 9 из 16 (т.е. 9/16) теоретически ожидаемых потомков имеют одновременно два доминантных признака (желтые гладкие горошины — жг); 3/16 — доминантный и рецессивный признак (желтые морщинистые — жм); 3/16 — рецессивный и доминантный признаки (зеленые гладкие — зг) и 1/16 часть потомков имеет одновременно два рецессивных признака (зеленые и морщинистые — зм).
При анализе генотипов по решетке Пеннета мы обнаруживаем 9 различных классов в соотношении 1 : 2 : 2 : 4 : 1 : 2 : 1 : 2 : 1.
Расщепление в дигибридном скрещивании в F![]() по фенотипу и генотипу можно получить, перемножая относительные частоты отдельных фенотипов или отдельных генотипов, поскольку гены, контролирующие развитие различных признаков, наследовались независимо друг от друга. По каждому из независимых признаков (окраска горошин и характеристика их поверхности) отношение частот, как было ранее показано, составляет 3:1. Тогда, перемножая (Зж:1з) на (Зг:1м), получаем 9жг : 3жм : 3зг : 1зм, что точно соответствует данным решетки Пеннета.
по фенотипу и генотипу можно получить, перемножая относительные частоты отдельных фенотипов или отдельных генотипов, поскольку гены, контролирующие развитие различных признаков, наследовались независимо друг от друга. По каждому из независимых признаков (окраска горошин и характеристика их поверхности) отношение частот, как было ранее показано, составляет 3:1. Тогда, перемножая (Зж:1з) на (Зг:1м), получаем 9жг : 3жм : 3зг : 1зм, что точно соответствует данным решетки Пеннета.
Рис. V.4. Схема дигибридного скрещивания.
Гибриды первого поколения единообразны как по фенотипу, так и по генотипу. Они образуют гаметы четырех различных типов — АВ, Ab, aB, ab
Таблица V.2
Расщепление по фенотипу и генотипу в F2 при дигибридном скрещивании:
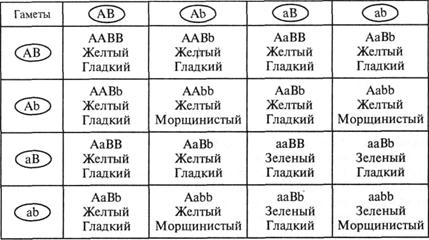
Таблица V.3
Соотношение фенотипических и генотипических классов во втором поколении при моно-, ди-, три- и полигибридном скрещивании:
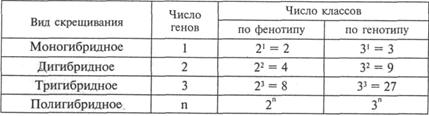

Зная, что при моногибридном скрещивании расщепление по генотипу соответствует 1АА: 2Аа: 1аа для одной пары и 1BB: 2Bb: 1bb для другой, можно подсчитать частоты, или вероятности, генотипов различных классов. Вероятности генотипов соответствуют: АА – ¼, Аа – ½, аа - ¼, ВВ – ¼, Вb – ½, bb – ¼. Например, относительная частота генотипа ААВВ рассчитывается путем перемножения вероятностей ¼АА х ¼BB = 1/16AABB, для ААВЬ — ¼AA х ½Вb = 1/8 или 2/16, ААВЬ. Тем же путем получаем распределение всех остальных различающихся по генетической конституции классов особей в отношении 1 : 2 : 2 : 4 : 1 : 2 : 1 : 2 : 1, что также полностью соответствует данным решетки Пеннета.
Поступая аналогичным образом, можно представить результаты расщепления по фенотипу и генотипу для тригибридного скрещивания, когда родительские формы различаются по трем независимым признакам и в F![]() , образуются тригибриды. Эксперименты показывают, что при тригибридном скрещивании расщепление в F
, образуются тригибриды. Эксперименты показывают, что при тригибридном скрещивании расщепление в F![]() по фенотипу дает 8 различных классов особей в соотношении 27:9:9:9:3:3:3:1, а расщепление по генотипу дает 27 различных классов.
по фенотипу дает 8 различных классов особей в соотношении 27:9:9:9:3:3:3:1, а расщепление по генотипу дает 27 различных классов.
Подобным образом возможен расчет вероятностей фенотипических и генотипических классов для любого полигибридного скрещивания (табл. V.3).
В общем виде эти соотношения можно выразить простыми формулами: число фенотипических классов равно 2![]() , где «2» отражает парность аллелей, а показатель степени «n» — число независимых генов. Число генотипических классов равно З
, где «2» отражает парность аллелей, а показатель степени «n» — число независимых генов. Число генотипических классов равно З![]() , где основание степени «3» — число генотипических классов при моногибридном скрещивании, а показатель степени «n» — число генов.
, где основание степени «3» — число генотипических классов при моногибридном скрещивании, а показатель степени «n» — число генов.
Очевидно, что в основе приведенных формул лежат закономерности моногибридного скрещивания. Они справедливы для любого числа генов, но не превышающих гаплоидное число n.
Важно отметить, что закономерности, открытые Менделем, реализуются при анализе большого количества особей, поскольку малое количество в потомстве гибридов (например, дети одной семьи) может давать отклонения от точного соотношения ожидаемых классов расщепления в силу случайных событий.
Гибридологический анализ, разработанный Менделем, и результаты, полученные на его основе, заложили концепцию фундаментального понятия генетики и биологии в целом — понятие гена. В последние десятилетия XIXв. были обнаружены хромосомы, описаны митотическое и мейотическое деления клетки. Тем не менее не были известны материальные носители наследственной информации. Только после того как законы Менделя были открыты вновь в 1900г., сопоставление менделевского расщепления признаков и распределения хромосом в мейозе позволило сделать окончательный вывод о том, что именно хромосомы являются носителями генетической информации. Этими событиями ознаменовалось начало нового научного периода развития генетики, а наблюдения и выводы Менделя и в настоящее время составляют важнейшую главу учения о наследственности и изменчивости.
Взаимодействие генов
Описано несколько типов взаимодействия неаллельных генов: комплементарность, эпистаз и полимерия.
Комплементарность — взаимодействие неаллельных генов, которые обусловливают развитие нового признака, отсутствующего у родителей. Примером комплементарного действия у человека могут служить случаи, когда у глухих родителей рождаются дети с нормальным слухом.
Развитие нормального слуха находится под генетическим контролем десятков различных неаллельных генов, гомозиготное рецессивное состояние одного из которых может приводить к одной из форм наследственной глухоты. Таких форм у человека известно более 30. Если один из родителей является гомозиготой по рецессивному гену аа (рис. V.5), а другой — гомозиготой по другому рецессивному гену bb, то все их дети будут двойными гетерозиготами и, следовательно, слышащими, поскольку доминантные аллели будут взаимно дополнять друг друга (см. рис. V.5). Таким образом формируется новый по отношению к родителям признак — нормальный слух.
Эпистаз (от греч. epi — над + stasis — препятствие) — взаимодействие неаллельных генов, при котором наблюдается подавление проявления одного гена действием другого, неаллельного гена. Подавляющий ген называется геном-супрессором, а подавляемый — гипостатическим геном. По-видимому, действие гена-супрессора на подавляемый ген сходно с принципом доминантность — рецессивность. Но существенное различие заключается в том, что эти гены не являются аллельными, т.е. расположены в негомологичных хромосомах или занимают различные локусы в гомоло-
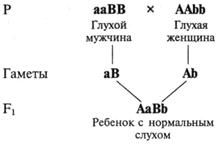
Рис. V.5. Схема, поясняющая возможность рождения детей с нормальным слухом у глухих родителей с различными генетическими формами глухоты.
гичных. Различают доминантный и рецессивный эпистаз. При доминантном эпистазе доминантный аллель гена-супрессора подавляет проявление доминантного аллеля другого гипостатического гена. При рецессивном эпистазе, или криптомерии, рецессивный аллель гена-супрессора, будучи в гомозиготном состоянии, не дает проявиться доминантной или рецессивным аллелям других генов.
Примером рецессивного эпистаза у человека может служить так называемый бомбейский фенотип, когда индивид, имеющий доминантный аллель группы крови системы АВ0 (например, аллель В, определяющий принадлежность человека к III или IV группе), идентифицируется в реакции агглютинации как человек с 0(I). Это состояние возникает в результате того, что данный индивид является рецессивной гомозиготой (hh) по другой, нежели система АВ0, генетической системе Hh. Для реализации аллелей I![]() и I
и I![]() необходимо присутствие хотя бы одного доминантного аллеля Н.
необходимо присутствие хотя бы одного доминантного аллеля Н.
Полимерия (от греч. polys — много + meros — часть) — вид взаимодействия, когда эффекты нескольких неаллельных генов, определяющих один и тот же признак, примерно одинаковы. Подобные признаки получили название количественных, или полимерных признаков. Как правило, степень проявления полимерных признаков зависит от числа доминантных генов. Наследование полимерных признаков было впервые описано шведским генетиком Г. Нильсон-Эле в 1908 г. Проводя скрещивание различных форм пшеницы (с красными и белыми зернами), он наблюдал расщепление в F![]() признака окраски в отношении: 15/16 окрашенных и 1/16 белых. Среди окрашенных зерен он наблюдал все переходы — от интенсивно окрашенных до слабо окрашенных.
признака окраски в отношении: 15/16 окрашенных и 1/16 белых. Среди окрашенных зерен он наблюдал все переходы — от интенсивно окрашенных до слабо окрашенных.
Анализ особенностей расщепления показал, что в данном случае окраску зерен определяют два доминантных аллеля двух различных генов, а сочетания их рецессивных аллелей определяют отсутствие окраски. Поскольку полимерные гены имеют однонаправленное действие, их, как правило, обозначают одинаковыми буквами. Таким образом, исходные родительские формы имели генотипы A![]() A
A![]() A
A![]() A
A![]() и a
и a![]() a
a![]() a
a![]() a
a![]() . Наличие всех четырех доминантных аллелей определяло самую интенсивную окраску, трех доминантных аллелей (типа A
. Наличие всех четырех доминантных аллелей определяло самую интенсивную окраску, трех доминантных аллелей (типа A![]() A
A![]() A
A![]() a
a![]() ) — менее интенсивную окраску и т.д.
) — менее интенсивную окраску и т.д.
Примером полимерного наследования у человека является наследование окраски кожных покровов. В браке индивида негроидной расы (коренного жителя Африки) с черной окраской кожи и представителем европеоидной расы с белой кожей дети рождаются с промежуточным цветом кожи (мулаты). В браке двух мулатов потомки могут обладать любой окраской кожи: от черной до белой, поскольку пигментация кожи обусловлена действием трех или четырех неаллельных генов. Влияние каждого из этих генов на окраску кожи примерно одинаково.
Полимерное наследование характерно для так называемых количественных признаков, таких, как рост, вес, окраска кожных покровов, скорость протекания биохимических реакций, артериальное давление, содержание сахара в крови, особенности нервной системы, уровень интеллекта, и многих других, которые нельзя разложить на четкие фенотипические классы. Чем большее число неаллельных генов контролируют развитие количественного признака, тем менее заметны переходы между фенотипическими классами.
Неменделевская генетика.
Гениальность законов Менделя заключается в их простоте. Строгая и элегантная модель, построенная на основе этих законов, служила генетикам точкой отчета на протяжении многих лет. Однако в ходе дальнейших исследований выяснилось, что законам Менделя подчиняются только относительно немногие генетически контролируемые признаки. Оказалось, что у человека большинство и нормальных, и патологических признаков детерминируются иными генетическими механизмами, которые стали обозначать термином «неменделевская генетика». Таких механизмов существует множество: хромосомные аберрации (синдром Дауна); наследование, сцепленное с полом (цветовая слепота); импринтинг (синдромы Прадера—Вилли, Энгельмана); появление новых мутаций (развитие раковых заболеваний); экспансия (инсерция) повторяющихся нуклеотидных последовательностей (миотоническая дистрофия); наследование количественных признаков (сложные поведенческие характеристики).
Хромосомные аберрации: синдром Дауна
Синдром Дауна (СД) - одно из весьма ограниченного числа наследуемых заболеваний, фенотип которого хорошо известен даже неспециалистам. Его «известность» является результатом того, что, во-первых, частота встречаемости СД достаточно высока и, во-вторых, фенотип этого заболевания легко узнаваем: больным СД свойственны характерные внешние черты, выражение лица и умственная отсталость.
Первые клинические и научные описания СД появились в середине прошлого века, а его точное определение было дано в 1866 г. Дж. Дауном, описавшим несколько таких пациентов. Гипотезы о том, что СД контролируется генетически, были сформулированы в начале XX в. К 30-м годам было высказано предположение, что это заболевание развивается в результате аберрации хромосом (структурных отклонений в хромосомном наборе), причиной которой служит их нерасхождение в процессе мейоза. В 1959 г. было обнаружено, что СД вызывается трисомией хромосомы 21, т.е. наличием в клетках трех, а не двух, как обычно, хромосом. Сегодня известно, что примерно 1 из 600 новорожденных является носителем этой аномалии. Кроме того, по современным оценкам, примерно 1 из 150 оплодотворенных яйцеклеток человека является носительницей трисомии 21 (большинство яйцеклеток с трисомиями гибнет). Пациенты с СД составляют около 25% всех умственно отсталых, формируя самую большую этиологически однородную группу умственно отсталых.
Генетический механизм СД представляет собой иллюстрацию явления хромосомных аберраций. Во время формирования половых клеток — гамет — все 23 пары хромосом делятся, и каждая гамета становится носителем одной хромосомы из каждой пары. Когда спермий оплодотворяет яйцеклетку, хромосомные пары восстанавливаются, причем в каждой паре одна хромосома приходит от матери, вторая — от отца. Несмотря на отлаженность процесса образования гамет, в нем случаются ошибки, и тогда разделение хромосомных пар нарушается — появляется гамета, которая содержит не одну хромосому, а их пару. Это нарушение называется нерасхождением хромосом. Когда такая гамета при оплодотворении сливается с нормальной гаметой, образуется клетка с тремя одинаковыми хромосомами; подобное явление и называется трисомией (см. рис. 1.7). Нерасхождение хромосом служит главной причиной спонтанных абортов в течение первых нескольких недель жизни плода. Тем не менее существует некоторая вероятность того, что зародыш с аномальным хромосомным набором продолжит развитие.
Точная причина нерасхождения неизвестна. Надежным коррелятом трисомии-21 является возраст матери: согласно исследованиям, у 56% матерей старше 35 лет плоды оказываются носителями трисомии-21, и в таких случаях примерно 90% диагностированных женщин предпочитают искусственно прервать беременность. Поскольку СД появляется «заново» в каждом поколении (нерасхождение — единичное событие, вероятность появления которого увеличивается с возрастом матери), постольку СД нельзя рассматривать как заболевание, передающееся по наследству.
Наследование, сцепленное с полом (х-хромосомой): цветовая слепота.
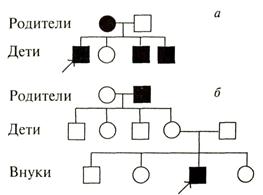
Рис. 3.1. Родословные пробандов, страдающих цветовой слепотой (признак наследуется по поло-сцепленному ме