Клетка: строение и внутренние процессы
Вопрос 1. Транскрипция. Структура и функции ядерного
хроматина
В зависимости от выполняемых функций РНК бывают несколько видов: и-РНК – информационная или матричная РНК – переносит информацию о структуре белка от ДНК к рибосомам, она составляет 1% от общего содержания;т-РНК (транспортная) переносит аминокислоты из цитоплазмы в рибосомы, на долю т-РНК приходится около 10% от общего количества РНК; р-РНК (рибосомальная) – составляет одну из субъединиц рибосомы, на ее долю приходится около 90% от всех РНК клетки.
ДНК – носитель генетической информации. В настоящее время геном называют участок молекулы ДНК, кодирующий первичную структуру полипептида. ДНК непосредственного участия в синтезе белков не принимает. Информацию несет посредник – и-РНК, который по принципу комплиментарности считывает (копирует) с ДНК информацию при участии фермента РНК-полимеразы. Переписывание последовательности нуклеотидов или генетической информации происходит с одной нити ДНК и называется транскрипцией (лат. transcriptio – переписывание). По длине каждая из молекул и-РНК в сотни раз короче ДНК. Информационная РНК является копией не всей молекулы ДНК, а только ее части – одного гена, несущего информацию о структуре белка. Готовая и-РНК отходит от ДНК и направляется к месту синтеза белка. В качестве матрицы используется молекула нуклеиновой кислоты.
Молекулярная сущность транскрипции выглядит так: Рис. 1 (В. Процесс транскрипции).
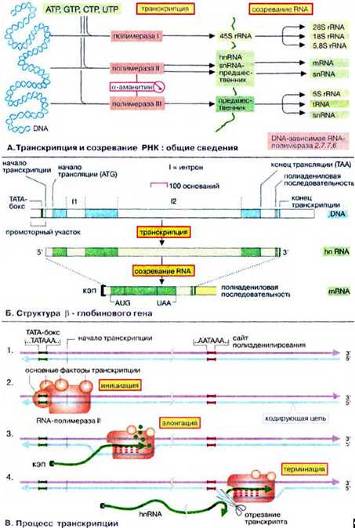
Рис. 1. – В. Процесс транскрипции
Процессинг РНК. Первоначальный «транскрипт» с ДНК значительно больше, чем зрелая РНК. Наряду со «значащими» участниками рибонуклеотидной последовательности транскрипта, так называемыми «экзонами», которые войдут в готовую молекулу РНК, в нем имеются и лишние, «молчащие» участки – «интроны», подлежащие удалению. Все интроны транскрибируются в составе РНК-предшественника и впоследствии удаляются в процессе разрыва-воссоединения – сплайсинга. Сплайсинг происходит еще в ядре, перед выходом РНК в цитоплазму.
Почти вся ДНК эукариотической клетки сосредоточена в ядре. Ядро окружено ядерной оболочкой, состоящей из двух концентрически расположенных мембран. Ядерные мембраны пронизаны удаленными на некоторое расстояние друг от друга порами, которые играют важную роль в переносе определенных молекул в цитоплазму из нее. Ядерная оболочка непосредственно связана с эндоплазматическим ретикулумом. С обеих сторон к ней прилегают сетеподобные структуры, состоящие из промежуточных филаментов. Та из них, которая выстилает внутреннюю ядерную мембрану, имеет вид тонкой оболочки и называется ядерной ламиной. Общий рисунок клетки (Рис. 2.), ядра (Рис. 3).
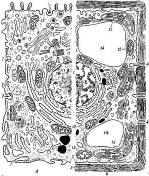
Рис. 2. – Общий рисунок клетки
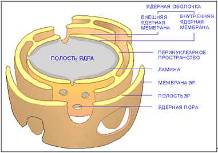
Рис. 3. – Ядро
Комбинированная схема строения эукариотпческой клетки.
А – клетка животного происхождения; В – растительная клетка: 1 – ядро с хроматином и ядрышком; 2 – клеточная (плазматическая) мембрана; 3 – клеточная оболочка; 4–плазмодесмы; 5 – гранулярная эндоплазматическая сеть; 6 – гладкая (агранулярная) эндоплазматическая сеть; 7 – пиноцитозная вакуоль; 8 – комплекс Гольджи; 9 – лизосома; 10 – жировые включения в гладкой эндоплазматической сети; 11 – центриоль и микротрубочки центра сферы; 12 – митохондрии; 13 – полирибосомы гиалоплазмы; 14 – вакуоли; 15 – хлоропласты.
ХРОМАТИН, вещество (нуклеопротеид) клеточного ядра, в процессе клеточного деления конденсируется, образуя компактные структуры – хромосомы. В состав хроматина входят: ДНК (30-40% по массе), гистоны (30-50%), негистоновые белки (4-33%) и РНК. Было исследовано регуляторное действие гистонов и негистоновых хромосомных белков. Как выяснилось, гистоны, оказывают тормозящее действие на ДНК-зависимый синтез РНК. Негистоновым хромосомным белкам тоже приписывают специфические регуляторные функции. Эти белки снимают блокирующее действие гистонов. Для хроматина известны 4 уровня укладки в более сложные структуры:нуклеосомы, нуклеомеры (или соленоид), хромомеры (или розетки с петельными доменами), хроматида (или хромосома). Различают эухроматин и гетерохроматин. Гетерохроматин, вещество хромосом, сохраняющее компактную (спирализованную) структуру на всех стадиях клеточного цикла. Эухроматин сохраняет деспирализованное (диффузное) состояние в покоящемся ядре и спирализующееся при делении клеток.
Таким образом, регуляция транскрипции – процесса переноса генетической информации от ДНК к РНК очень нужна и важна с позиций устройства многоклеточного организма.
Вопрос 2. Организация пластического метаболизма на примере
секреторной клетки
Пластический метаболизм (обмен веществ) – это превращения разл. соед. в организме, их перенос (транспорт) внутри организма и между организмом и средой.
Биосинтез белков начинается на рибосомах. Однако вскоре пути синтезируемых белков расходятся в соответствии с их функцией: белки, несущие на N-конце сигнальный пептид для ЭР, проходят через секреторный путь, а прочие белки, не имеющие этой сигнальной последовательности, следуют по цитоплазматическому пути. Рис. 4. (А. Транспорт белков).
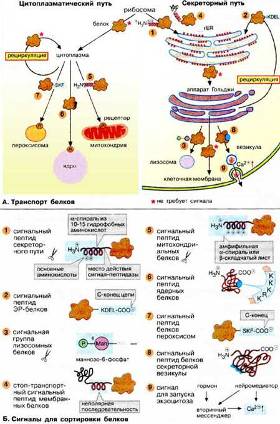
Рис. 4. – А. Транспорт белков
Трансляция – процесс синтеза белка в цитоплазме клетки. Началом принято считать процесс транскрипции ДНК. Процесс трансляции начинается с присоединения малой рибосомной субчастицы к молекуле мРНК. Особая инициаторная т-РНК связывает малую рибосомную субчастицу со специальным старт- кодоном на м-РНК. Присоединение большой субчастицы завершает сборку рибосомы. Далее следует фаза элонгации: связывания аминоацил-тРНК, образования пептидной связи и транслокации рибосомы. К стоп-кодону присоединяется затем фактор освобождения, останавливающий трансляцию и вызывающий отделение завершенного полипептида от рибосомы.
Рибосомы состоят из 2 различных субчастиц, каждая из которых построена из рибосомнойРНК и многих белков. Рис. 5 (А.Структура рибосомэукариот).
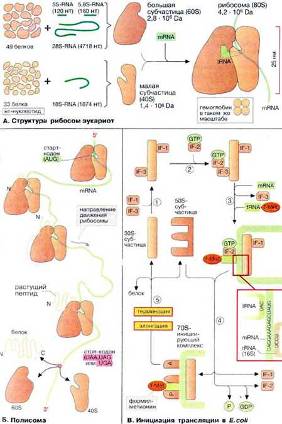
Рис. 5. – А.Структура рибосомэукариот
Все эукариотические клетки имеют эндоплазматический ретикулум (ЭР): с цистернами шероховатого ЭР соединены мембраны гладкого ЭР. Шероховатый ЭР усеян рибосомами, расположенными на его цитоплазматической поверхности; он образует поляризованные стопки уплощенных цистерн.
Функция ШЭР: активный биосинтез белков. Рис. 6.
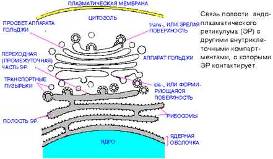
Рис. 6. – ЭР
Аппарат Гольджи управляет процессами внутриклеточного транспорта. Основными его функциями являются модификация, накопление, сортировка и направление различных веществ в соответствующие внутриклеточные компартменты, а также за пределы клетки. Он состоит из набора окруженных мембраной уплощенных цистерн, напоминающих стопку тарелок. Аппарат Гольджи имеет две разные стороны: формирующуюся (цис-сторону) и зрелую (транс-сторону). Белки и липиды в составе небольших пузырьков попадают в стопку Гольджи с цис-стороны, а покидают ее, направляясь в различные компартменты, вместе с пузырьками, образующимися на транс-стороне. Решающая роль аппарата Гольджи: регенерация и обновление своей мембраны.
Лизосомы содержат пищеварительные ферменты, которые разрушают отработанные органеллы, а также частицы и молекулы, поглощенные клеткой извне путем эндоцитоза. Рис. 7 (Б).
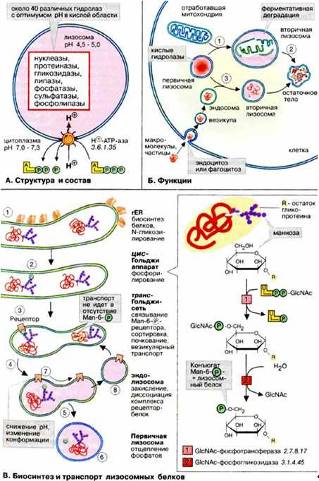
Рис. 7 (Б)
Вопрос 3. Энергетический метаболизм и использование энергии
В энергетич. обмене рассматривают превращения хим. энергии, образующейся в обмене веществ, в тепло, мышечную работу, а также механизмы ее использования в активном транспорте, биосинтезе и др.
В клетке химическая энергия запасается в виде так называемых «высокоэнергетических» метаболитов. Наиболее важным таким метаболитом, макроэргом обеспечивающим энергией большое число энергозависимых реакций, является АТФ. Рис. 8 (А).
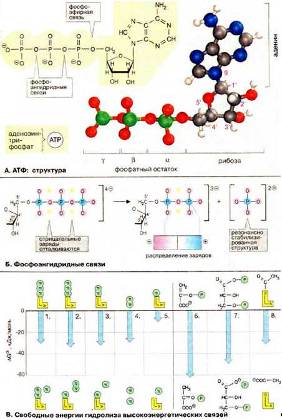
Рис. 8 (А)
АТФ = АДФ + Ф.
Суть этого цикла: При гидролизе концевой фосфоангидридной связи АТФ превращается в АДФ и ортофосфат, и освобождается большое количество свободной энергии. Величина свободной энергии гидролиза АТФ делает возможным его образование из АДФ за счет переноса фосфатного остатка от других высокоэнергетических фосфатов.
У животных, растений и грибов синтез АТФ протекает в специализир. субклеточных структурах-митохондриях. В зеленых водорослях и высших растениях фотосинтез происходит в хлоропластах.
Энергия света в процессе фотосинтеза используется для синтеза органических соединений из CO2 и воды. В световой реакции образуется АТФ.
![]()
Гликолиз – это катаболический путь обмена веществ в цитоплазме. Баланс гликолиза простой: в аэробных условиях молекула глюкозы деградирует до двух молекул пирувата. Кроме того, образуются по две молекулы АТФ и НАДН + H+ (аэробныйгликолиз). В анаэробных условиях пируват претерпевает дальнейшие превращения, обеспечивая при этом регенерацию НАД+. При этом образуются продукты брожения, такие, как лактат или этанол (анаэробныйгликолиз).
![]()
Окислительное фосфорилирование, синтез АТФ из аденозиндифосфата и неорг. фосфата, осуществляющийся в живых клетках, благодаря энергии, выделяющейся при окислении орг. в-в в процессе клеточного дыхания. В общем виде окислительное фосфорилирование и его место в обмене в-в можно представить схемой:
Митохондрии и хлоропласты осуществляют синтез АТФ. Рис. 9., согласно которому: а) – митохондрии (указаны стрелкой), видимые в световом микроскопе; б – ультраструктура митохондрий: 1 – митохондриальный матрикс, 2 – внутренняя митохондриальная мембрана, 3 – межмембранное пространство, 4 – внешняя митохондриальная мембрана; в – общая схема функционирования митохондрий: при переносе электронов в цепи окисления в межмембранном пространстве накапливаются протоны и при достижении определенного потенциала возвращаются в матрикс; энергия этого потенциала тратится на синтез АТФ.
Рис. 9
Хлоропласты – это органеллы, которые, подобно митохондриям, окружены двумя мембранами. Во внутреннем пространстве, строме, находятся тилакоиды, уплощенные мембранные мешки, которые будучи сложены стопками образуют граны. Внутреннее содержимое тилакоида называют люменом. Рис. 10 (в голубой рамке).
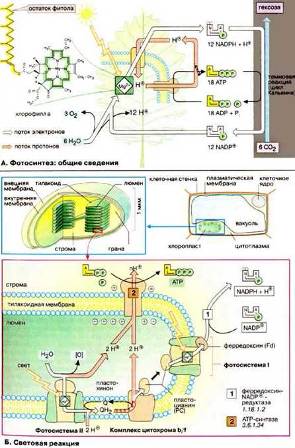
Рис. 10 (в голубой рамке)
Вопрос 4. Опорно-двигательная система клетки (Цитоскелет)
ядерный метаболизм клетка мембрана
Цитоскелет – это совокупность фибриллярных компонентов цитоплазмы эукариотических клеток. Основными типами фибрилл в составе цитоскелета являются актиновые филаменты, микротрубочки и промежуточные филаменты. Относительно функций промежуточных филаментов известно очень мало. Предполагается, что промежуточные филаменты играют механическую роль в межклеточных взаимодействиях и в организации тканевой структуры. В состав промежуточных филаментов у клеток из различных тканей входят следующие белки: виментин, десмин, глиальный фибриллярный кислый белок, белки нейрофиламентов, кератины, ламины.
Схема строения актиновых микрофиламентов на Рис. 11 (А).
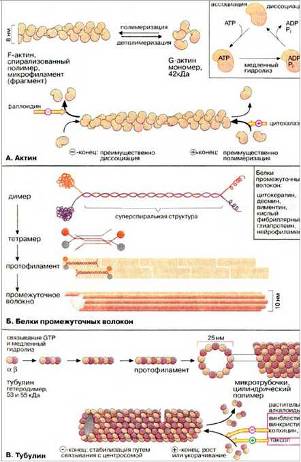
Рис. 11 (А)
Актиновые филаменты или фибриллярный актин (F-актин) представляют собой тонкие фибриллы диаметром 6-8 нм. Они являются результатом полимеризации глобулярного актина – G-актина. Актиновые филаменты играют ключевую роль в сократительном аппарате мышечных и немышечных клеток, а также принимают участие во многих других клеточных процессах, таких как подвижность, поддержание формы клеток, цитокинез. Существуют системы в которых движение органелл происходит по актиновым филаментам. Движение органелл по актиновым филаментам опосредуется миозинами (актин-связывающими белками), которые объединяются в одну группу благодаря наличию общего "головного" домена, обладающего АТФазной активностью.
Схема строения микротрубочки на Рис. 12.
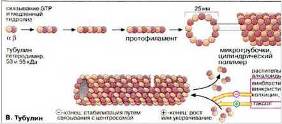
Рис. 12
Белком микротрубочек является тубулин, полимеризация которого приводит к образованию этих структур. Еще есть белки, ассоциированные с микротрубочками (MAP) и белки – транслокаторы. Тубулин – очень консервативный белок, связывающий две молекулы ГТФ, и может также связывать ионы кальция и магния. Функция динеина в биении ресничек и жгутиков, преобразуя энергию АТР в механическое усилие. Микротрубочки имеют две основные функции – двигательную и структурную. Двигательная функция заключается в том, что по микротрубочкам с помощью специальных транспортных белков – транслокаторов – осуществляется движение клеточных органелл. Структурная функция состоит в поддержании с помощью микротрубочек определенной формы клетки или ее части.
В клетках концы микротрубочек, как правило, ассоциированы со специальными структурами – ЦОМТ (центр организации микротрубочек). Некоторые ЦОМТ (например, центриоли, базальные тельца) содержат систему сложно организованных микротрубочек. Другие же представляют собой скопления аморфного электронно-плотного материала. Эти скопления могут быть ассоциированы с другими клеточными компонентами, такими как центриоли, базальные тельца, ядерная оболочка, плазматическая мембрана. Типы ЦOМТ: центросомы, базальные тельца, кинетохоры. Рис. 13 (Б, кружочек).
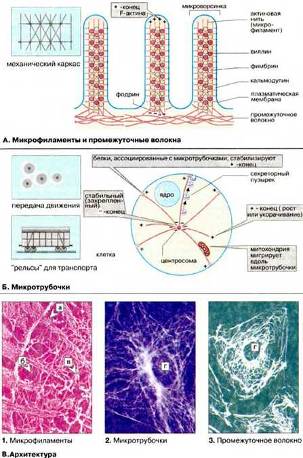
Рис. 13 (Б, кружочек)
В большинстве животных клеток область, где впервые образуется веретено, содержит центриоли. Каждая пара центриолей в митозе становится частью митотического центра, от которого лучами расходятся микротрубочки. Между тем у многих организмов, в том числе у высших растений, функционально полноценное веретено образуется при полном отсутствии центриолей. Кроме того, если у живой клетки разрушить лазерным микролучом центриоли, то митотическое веретено продолжает нормально функционировать. По-видимому, центриоли не являются структурами, необходимыми для сборки микротрубочек веретена; но если центриоли в клетке есть, то они играют роль фокусов, в которых сходятся микротрубочки.
Вопрос 5. Поверхностный аппарат клетки
Плазматическая мембрана, окружающая каждую клетку, определяет ее величину и обеспечивает сохранение существенных различий между клеточным содержимым и окружающей средой. Мембрана служит высокоизбирательным фильтром, который поддерживает разницу концентраций ионов по обе стороны мембраны и позволяет питательным веществам проникать внутрь клетки, а продуктам выделения выходить наружу.
Жидкостно-мозаичная модель строения плазматической мембраны на Рис. 14 (А).
Пассивный транспорт – это прохождение растворенных веществ через липидный бислой посредством электрохимического градиента.
Активный транспорт – это перекачка растворенных веществ против их электрохимических градиентов белками-переносчиками. Активный транспорт обязательно связан с источником метаболической энергии. Чаще всего таким источником служит гидролиз АТР, осуществляемый белками-переносчиками, или котранспорт ионов натрия или котранспорт ионов водорода по их электрохимическим градиентам.
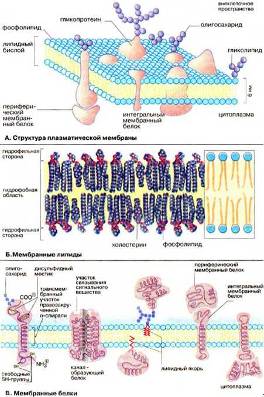
Рис. 14 (А)
Растворенные вещества способны так же перемещаться через саму мембрану – диффузия.
Кроме того, Активный транспорт может идти по механизму унипорта (облегченной диффузии), согласно которому только одно вещество переносится через биомембрану в одном направлении с помощью канальных или транспортных белков (например, транспорт глюкозы в клетках печени). Активный транспорт может протекать по механизму сопряженного переноса (симпорт, сопряженный транспорт), когда два вещества переносятся одновременно в одном направлении как например, транспорт аминокислот или глюкозы вместе с ионами натрия в кишечных эпителиальных клетках, либо в противоположном направлении (антипорт, обменная диффузия), как, например, обмен ионов НСО3- на Cl- в мембране эритроцитов.
Высокое соотношение концентрации калия во вне- и внутриклеточной жидкости (38:1) поддерживается благодаря действию Nа+,К+-АТФазы, активно переносящей ионы калия в клетку, а ионы натрия – из нее (в соотношении 2:3) (вследствие активного выведения натрия из клеток Na+,K+-АТФазой 85-90% всего натрия, содержащегося в организме, находится во внеклеточной жидкости и по этой причине определяет ее объем).
Межклеточные контакты возникают в местах соприкосновения клеток в тканях и служат для межклеточного транспорта веществ и передачи сигналов, а также для механического скрепления клеток друг с другом. Типы:
а) рыхлые (простые) контакты – между плазматическими мембранами соседних клеток имеется щель шириной 10-20 нм, заполненная гликокалликсом, специализированных структур на мембранах нет;
б) межклеточные «замки» – мембраны соседних клеток разделены таким же расстоянием, но изгибаются, образуя на поверхности клеток впячивания;
в) десмосомы;
г) плотные контакты (встречаются в основном в эпителиальных клетках) – разделяются на зону замыкания и зону слипания (промежуточный контакт); в зоне замыкания две соседние мембраны сливаются своими наружными слоями, эта зона непроницаема для макромолекул и ионов, в зоне слипания мембраны разделены щелью в 10-20 нм, заполненной плотным веществом, вероятно, белковой природы;
д) щелевидные (высокопроницаемые) контакты, свойственные всем типам эпителиальной и соединительной тканей, – плазматической мембраны разделены промежутком в 2-4 нм, пронизанным каналами, по которым низкомолекулярные вещества попадают из цитоплазмы одной клетки в другую, минуя межклеточную среду.
Гликокаликс – это обогащенная углеводами периферическая зона на поверхности большинства эукариотических клеток (клеточная оболочка). Гликокаликс выполняет прежде всего функцию непосредственной связи клеток животных с внешней средой, со всеми окружающими ее веществами.
1. Биология. (Учебник) / Под ред. В.Н. Ярыгина. – 5-е изд. – 2003.
2. Микробиология. (Учебник) / Гусев М.В., Минеева Л.А. – 2003.
3. Пехов А.П. Биология с основами экологии. (Учебник). – 2000.
4. Медицинская генетика / Под ред. Бочкова Н.П. – М.: Мастерство, 2001.
5. Ярыгин В.Н., Волков И.Н. и др. Биология. – М.: Владос, 2001.
6. Биология / Под ред. Чебышева. Н.В. – М.: ГОУ ВУНМЦ, 2005.
7. Орехова. В.А., Лажковская Т.А., Шейбак М.П. Медицинская генетика. – Минск: Высшая школа, 1999.
8. Пособие по биологии для довузовского обучения иностранных учащихся / Под ред. Чернышова В.Н., Елизаровой Л.Ю., Шведовой Л.П. – М.: ГОУ ВУНМЦ МЗ РФ, 2004.
9. Врожденные пороки развития // Серия учебной литературы «Образование медсестер», модуль 10. – М.: Гэотар-мед, 2002.