Цитология, эмбриология, общая гистология
Луганский национальный аграрный университет
Цитология, эмбриология, общая гистология
(курс лекций)
Луганск - 2005
Цитология, эмбриология, общая гистология
Курс лекций составлен заведующим кафедрой биологии животных, доктором биологических наук, профессором Г.Д. Кацы.
Издание 2-е, переработанное и дополненное.
Лекции подготовлены для студентов зообиотехнологического и факультета ветеринарной медицины Луганского национального аграрного университета. Искренне благодарю аспиранта кафедры биологии животных Крыцю Я.П. и заведующую лабораторией Есауленко В.П. за помощь при подготовке материала к изданию.
Введение в гистологию
1. Предмет гистологии и её место в системе биологических и ветеринарных наук.
2. История и методы микроскопических исследований.
3. Клеточная теория, основные положения.
1. Специфика сельскохозяйственного производства обусловлена тем: что несмотря на возрастание роли технических факторов: главными орудиями и средствами производства остаются биологические объекты. По охвату объектов изучения и по своей глубине ветеринария представляет: как говорил академик К.И.Скрябин, интереснейшую область человеческого знания: в которой исследуется и охраняется такое множество представителей животного царства.
Цитология, гистология и эмбриология, наряду с физиологией, биохимией и другими науками формирует фундамент современной ветеринарии.
Гистология (греч. histos-ткань, logos-учение)- наука о развитии, строении и жизнедеятельности тканей животных организмов. Современная гистология изучает структуры организма животных и человека в связи с происходящими в них процессами, раскрывает соотношения между функцией и структурой и т.д.
Гистологию делят на 3 основных раздела: цитологию, или учение о клетке; эмбриологию, или учение о зародыше и гистологию общую и частную, или учение о тканях, о микроскопическом строении органов, их клеточном и тканевом составе.
Гистология тесно связана с рядом биологических и ветеринарных наук - общей и сравнительной анатомией, физиологией, патологической физиологией и патологической анатомией, а также некоторыми клиническими дисциплинами (внутренние болезни, акушерство и гинекология и др.).
Будущим врачам необходимо хорошее знание строения клеток и тканей органов, являющихся структурной основой всех видов жизнедеятельности организма. Значимость гистологии, цитологии и эмбриологии для врачей возрастает ещё потому, что для современной ветеринарной медицины характерно широкое применение цитологических и гистологических методов при проведении анализов крови, костного мозга, биопсии органов и пр.
2. Понятие ткань впервые было введено в биологию блестящим молодым французским ученым анатомом и физиологом Ксавье Биша (Bichat, 1771-1802), на которого произвела такое сильное впечатление разнообразная текстура различных слоев и структур, обнаруженных им при анатомических исследованиях, что он написал книгу о тканях организма, дав в ней название более чем 20 их видам.
Термин “”гистология” не принадлежит Биша, хотя его и можно считать первым гистологом. Термин “гистология” через 17 лет после смерти Биша предложил немецкий исследователь Мейер.
Ткань есть филогенетически обусловленная элементарная система, объединенная общей стуктурой, функцией и развитием (А.А. Заварзин).
Успехи гистологии с момента зарождения и по настоящее время прежде всего связаны с развитием техники, оптики и методов микроскопирования. Историю гистологии можно разделить на три периода: 1-й - домикроскопический (продолжительность около 2000 лет), 2-й - микроскопический (около 300 лет), 3-й - электронно-микроскопический (около 40 лет).
В современной гистологии, цитологии и эмбриологии применяются разнообразные методы исследования, позволяющие всесторонне изучать процессы развития, строения и функции клеток, тканей и органов.
Объектами исследования служат живые и мертвые (фиксированные) клетки и ткани, их изображения, полученные в световых и электронных микроскопах или на телевизионном экране. Существует ряд методов, позволяющих проводить анализ указанных объектов:
1) методы исследования живых клеток и тканей: а) прижизненное исследование клеток в организме (in vivo) - с помощью методов вживления прозрачных камер в организм животных, методом трансплантации;
б) исследование живых структур в культуре клеток и тканей (in vitro) - недостатки: утрачивается взаимосвязь с другими клетками и тканями, действие комплекса нейрогуморальных факторов регуляции и другое;
в) витальное и суправитальное окрашивание, то есть прижизненное окрашивание и окрашивание живых клеток, выделенных из организма.
2) исследование мертвых клеток и тканей; основным объектом исследования здесь являются гистологические препараты, приготовляемые из фиксированных структур.
Процесс изготовления гистопрепарата для световой и электронной микроскопии включает следующие основные этапы: 1) взятие материала и его фиксация, 2) уплотнение материала, 3) приготовление срезов, 4) окрашивание или контрастирование цвета. Для световой микроскопии необходим ешё один этап - заключение срезов в бальзам или другие прозрачные среды (5).
3) исследование химического состава и метаболизма клеток и тканей:
- цито- и гистохимические методы,
- метод радиоавтографии, в основе которого лежит использование радиоактивных элементов (например, фосфора-32Р, углерода -14С, серы-35S, водорода-3Н) или меченных им соединений.
- метод дифференциального центрифугирования - метод основан на применении центрифуг, дающих от 20 до 150 тыс. оборотов в минуту. При этом отделяются и осаждаются различные компоненты клеток и определяется их химический состав. - интерферометрия - метод позволяет оценить сухую массу и концентрацию плотных веществ в живой и фиксированной клетках. - количественные гистохимические методы - цитоспектрофотометрия - метод количественного изучения внутриклеточных веществ по их абсорбционным свойствам. Цитоспектрофлюориметрия - метод изучения внутриклеточных веществ по спектрам их флюорисценции.
4) методы иммунофлюорисцентного анализа. Они применяются для изучения процессов дифференцировки клеток, выявления в них специфических химических соединений и структур. Они основаны на реакциях антиген-антитело.
Методы микроскопирования гистологических препаратов:
- световая микроскопия: а) ультрафиолетовая, б) флюоресцентная (люминисцентная).
- электронная микроскопия: а) просвечивающая, б) сканирование (считывание). Первая дает лишь плоскостное изображение, вторая - пространственное; главным достоинством последнего (растрового) является большая глубина резкости (в 100-1000 раз больше, чем у световых микроскопов), широкий диапазон непрерывного изменения увеличения (от десятков до десятков тысяч раз) и высокая разрешающая способность.
3. Организм высших животных состоит из микроскопических элементов - клеток и ряда их производных - волокон, аморфного вещества.
Значение клетки в многоклеточном организме определяется тем, что через неё передается наследственная информация, с неё начинается развитие многоклеточных животных; благодаря деятельности клеток образуются неклеточные структуры и основное вещество, которые вместе с клетками образуют ткани и органы, выполняющие специфические функции в сложном организме. Создателем клеточной теории следует считать Дютроше (1824, 1837) и Шванна (1839).
Дютроше (1776-1847) - зоолог, ботаник, морфолог, физиолог. В 1824 г. он опубликовал свою книгу “”Анатомические и физиологические исследования о тонком строении животных и растений, а также о их подвижности”.
Созданию клеточной теории предшествовали следующие открытия. В 1610 году 46-летний проф. матетатики Падуанского университета Г.Галилей сконструировал микроскоп. В 1665 г. Роберт Гук открыл клетку при увеличении 100 х. Его современник, Феличе Фонтана говорил: “”...Посмотреть в микроскоп может каждый, но лишь немногие могут судить о виденном”. “”Микрография” Гука включала 54 наблюдения, в т.ч.”Наблюдение 18. О схематизме или строении пробки или о клетках и порах в некоторых других рыхлых телах”.
Из истории. Компания живших в Лондоне молодых людей (студентов) в 1645 г. стала собираться каждый день после занятий, чтобы обсуждать проблемы экспериментальной философии. Среди них были Роберт Бойль (18 лет), Р.Гук (17 лет), Рэн (23 года) и др. Так зародилась Британская академия, затем Лондонское Королевское общество (Карл II был её почетным членом).
Животную клетку открыл Антон ван Левенгук (1673-1695). Жил он в Делфте и торговал сукном. Свои микроскопы довел до 275 х. Петру I показывал кровообращение в хвосте личинки угря.
В настоящее время клеточная теория гласит: 1) клетка является наименьшей единицей живого, 2) клетки разных организмов сходны по своему строению, 3) размножение клеток происходит путем деления исходной клетки, 4) многоклеточные организмы представляют собой сложные ансамбли клеток и их производных, объединенные в целостные интегрированные системы тканей и органов, подчиненные и связанные между собой межклеточными, гуморальными и нервными формами регуляции.
Клетка - элементарная единица живого
1. Состав и физико-химические свойства живого вещества.
2. Типы клеток. Теории происхождения эукариотической клетки.
3. Клеточные мембраны, их молекулярный состав и функции.
1. Типичную клетку с ядром, цитоплазмой и всеми содержащимися в ней органеллами еще нельзя считать наименьшей единицей живого вещества, или протоплазмы (греч. “протос” -первый, “плазма” -образование). Существуют и более примитивные или более просто организованные единицы жизни - так называемые прокариотические организмы (греч. “карион” - ядро), к которым относится большинство вирусов, бактерии и некоторые водоросли; у них в отличие от клеток высшего типа с настоящим ядром (эукариотические клетки) отсутствует ядерная оболочка и ядерное вещество смешивается или непосредственно соприкасается с остальной протоплазмой.
В состав живого вещества входят белки, нуклеиновые кислоты (ДНК и РНК), полисахариды и липиды. Химические компоненты клетки можно разделить на неорганические (вода и минеральные соли) и органические (белки, углеводы, нуклеиновые кислоты, липиды и т.д.).
Цитоплазма растительной и животной клетки содержит 75-85 % воды, 10-20 % белка, 2-3 % липидов, 1 % углеводов и 1 % неорганических веществ.
ДНК - это молекула (её содержится 0,4 %), которая содержит генетическую информацию, направляющую синтез специфических клеточных белков. На одну молекулу ДНК приходится около 44 молекул РНК, 700 молекул белка и 7000 молекул липидов.
Первичная структура РНК подобна структуре ДНК, за исключением того, что РНК содержит рибозу и вместо тимина урацил. В настоящее время установлено, что существуют различающиеся молекулярным весом и другими свойствами три типа РНК: рибосомная, информационная и транспортная. Эти три типа РНК синтезируются в ядре и участвуют в синтезе белка.
2. Шаттон (1925) разделил все живые организмы на два типа (клистера) - прокариоты и эукариоты. Они дивергировали в докембрии (600-4500 млн. лет назад). Существуют две концепции происхождения эукариотической клетки: экзогенная (симбиотическая) и эндогенная. Первая основана на признании принципа объединения разных прокариотных организмов друг с другом. Эндогенная концепция основана на принципе прямой филиации, т.е. последовательного эволюционного преобразования прокариотных организмов в эукариотные.
В организме млекопитающих гистологи насчитывают около 150 типов клеток, и большинство из них приспособлено к выполнению какой-то одной определенной задачи. Форма и строение клетки зависят от выполняемой ею функции.
Функции клеток: раздражимость, сократимость, секреция, дыхание, проводимость, поглощение и усвоение, экскреция, рост и размножение.
3. Любую клетку отграничивает плазматическая мембрана. Она настолько тонка, что её невозможно различить под световым микроскопом. Плазматическая мембрана, легко поврежденная микроиглой, способна к восстановлению, но при более грубом повреждении, особенно в отсутствие ионов кальция, цитоплазма вытекает через прокол наружу и клетка погибает.
Согласно современной теории, плазматическая мембрана состоит из бислоя полярных липидов и встроенными в него молекулами глобулярных белков. Благодаря этим слоям мембрана, обладает эластичностью и относительной механической прочностью. Плазматическая мембрана большинства типов клеток состоит из трёх слоёв шириной примерно 2,5 нм каждый. Подобная структура, называемая “элементарной мембраной”, обнаружена и в большинстве внутриклеточных мембран. Биохимический анализ показал, что липиды и белки содержаться в них в отношении 1.0 : 1.7. Белковый компонент, названный строматином, представляет собой кислый фибриллярный белок с высоким молекулярным весом. Основную массу липидных компонентов образуют фосфолипиды, преимущественно лецитин и кефалин.
Плазмолемма - оболочка клетки, выполняющая отграничительную, транспортную и рецепторную функции. Она обеспечивает механическую связь клеток и межклеточные взаимодействия, содержит клеточные рецепторы гормонов и других сигналов окружающих клетку среды, осуществляет транспорт веществ в клетку из клетки как по градиенту концентраций - пассивный перенос, так и с затратами энергии против градиента концентраций - активный перенос.
В состав оболочки входят плазматическая мембрана, немембранный комплекс - гликокалекс и субмембранный опорно-сократительный аппарат.
В гликокалексе содержится около 1 % углеводов, молекулы которых образуют длинные ветвящиеся цепи полисахаридов, связанные с белками мембраны. Находящиеся в гликокалексе белки - ферменты участвуют в конечном внеклеточном расщеплении веществ. Продукты этих реакций в виде мономеров поступают в клетку. При активном переносе транспорт веществ в клетку осуществляется или поступлением молекул в виде раствора - пиноцитоз, или захватом крупных частиц - фагоцитоз.
В соответствии с функциональными и морфологическими особенностями тканей оболочка клеток образует характерные для них аппараты межклеточных контактов. Основные их формы: простой контакт (или зона слипания), плотный (замыкающий) и щелевой контакт. Разновидностью плотного контакта являются десмосомы.
Биологичекие мембраны действуют как диффузные барьеры. Благодаря своей избирательной проницаемости для ионов К+, Nа+, Cl- и т.п., а также высокомолекулярных соединений они разграничивают внутри- и межклеточные зоны реакций и создают электрические градиенты и градиенты концентрации веществ. Это делает возможным существование упорядоченных биологических структур со специфическими функциями.
Проникновение веществ в клетку называется эндоцитозом. Но существует и экзоцитоз. Например, от аппарата Гольджи отшнуровываются секреторные пузырьки, мигрирующие по направлению к клеточной мембране и выбрасывающие наружу своё содержимое. При этом мембрана пузырька сливается с гомологичной ей клеточной мембраной.
На основании электронно-микроскопических данных можно предположить, что плазмолемма является продуктом аппарата Гольджи. От этой органеллы в виде непрерывно отделяющихся пузырьков постоянно идет транспорт мембранного материала (“”поток мембран”), восстанавливающего использованные участки плазмолеммы и обеспечивающего её рост после деления клетки.
Мембрана является носителем видоспецифических и специфических для клетки поверхностных свойств, связанных с характерным распределением на ней гликозаминогликанов и белков. Их молекулы могут также покрывать поверхность клеток в виде тончайших пленок и образовывать межклеточный матрикс между соседними клетками. Контактные свойства клеток и иммунные реакции определяются этими компонентами мембран.
У многих клеток, особенно у специализированных для всасывания (кишечный эпителий), на наружной стороне имеются волосовидные выросты - микроворсинки. Образуемая или “щеточная каемка” несет ферменты, принимает участие в расщеплении веществ и транспортных процессах. На базальной стороне клеток, специализированных на интенсивное пропускание жидкости (при осморегуляции), например, в эпителии почечных канальцев и мальпигиевых сосудов, мембрана образует множественные впячивания, составляющие базальный лабиринт. Продукт клеточной секреции, базальная мембрана, часто отграничивает эпителий от глубже лежащих клеточных слоёв.
Особые мембранные структуры возникают в местах соприкосновения соседних клеток. Там имеются области, где мембраны так тесно прилегают друг к другу, что не остаётся места для межклеточного вещества (плотный контакт). В других участках возникают сложные контактные органеллы - десмосомы. Они и другие контактные структуры служат для механического соединения и главное - обеспечивают химическую и электрическую интеграцию соседних клеток, облегчая межклеточный ионный транспорт благодаря своему низкому электрическому сопротивлению.
Строение животной клетки
1. Цитоплазма и органеллы, их функция.
2. Ядро, его строение и функции.
3. Типы деления, фазы клеточного цикла.
1. Цитоплазма, отделенная от окружающей среды плазмолеммой, включает в себя гиалоплазму, находящиеся в ней обязательные клеточные компоненты - органеллы, а также различные непостоянные структуры – включения (рис.1).
Гиалоплазма (hyalinos - прозрачный) - основная плазма, или матрикс цитоплазмы, представляет собой очень важную часть клетки, её истинную внутреннюю среду.
В электронном микроскопе матрикс имеет вид гомогенного и тонкозернистого вещества с низкой электронной плотностью. Гиалоплазма является сложной коллоидной системой, включающей в себя различные биополимеры: белки, нуклеиновые кислоты, полисахариды и др. Эта система способна переходить из золеобразного (жидкого) состояния в гелеобразное и обратно. В состав гиалоплазмы входят главным образом различные глобулярные белки. Они составляют 20-25% общего содержания белков в эукариотической клетке. К важнейшим ферментам гиалоплазмы относятся ферменты метаболизма сахаров, азотистых оснований, аминокислот, липидов и других важных соединений. В гиалоплазме располагаются ферменты активации аминокислот при синтезе белков, транспортные РНК (тРНК). В гиалоплазме при участии рибосом и полирибосом происходит синтез белков, необходимых для собственно клеточных нужд, для поддержания и обеспечения жизни данной клетки.
Органеллы - постоянно присутствующие и обязательные для всех клеток микроструктуры, выполняющие жизненно важные функции.
Различают мембранные органеллы - митохондрии, эндоплазматическую сеть (гранулярная и гладкая), аппарат Гольджи, лизосомы, к категории мембранных органелл относится и плазмолемма; н е м ембранные органеллы: свободные рибосомы и полисомы, микротрубочки, центриоли и филаменты (микрофиламенты). Во многих клетках органеллы могут принимать участие в образовании особых структур, характерных для специализированных клеток. Так, реснички и жгутики образуются за счет центриолей и плазматической мембраны, микроворсинки - это выросты плазматической мембраны с гиалоплазмой и микрофиламентами, акросома спермиев - это производное элементов аппарата Гольджи и пр.
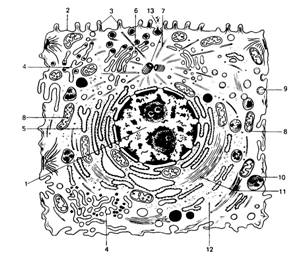
Рис 1. Ультрамикроскопическое строение клетки животных организмов (схема)
1 – ядро; 2 – плазмолемма; 3 – микроворсинки; 4 – агранулярная эндоплазматическая сеть; 5 - гранулярная эндоплазматическая сеть; 6 – аппарат Гольджи; 7 – центриоль и микротрубочки клеточного центра; 8 – митохондрии; 9 – цитоплазматические пузырьки; 10 – лизосомы; 11 – микрофиламенты; 12 – рибосомы; 13 – выделение гранул секрета.
Мембранные органеллы представляют собой одиночные или связанные друг с другом отсеки цитоплазмы, отграниченные мембраной от окружающей их гиалоплазмы, имеющие своё собственное содержимое, отличное по составу, свойствам и функциям:
Митохондрии - органеллы синтеза АТФ. Их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии для синтеза молекул АТФ. Митохондрии ещё называют энергетическими станциями клетки, или органеллами клеточного дыхания.
Термин “”митохондрия” был введён Бенда в 1897 году. Митохондрии можно наблюдать в живых клетках, т.к. они обладают достаточно высокой плотностью. В живых клетках митохондрии могут перемещаться, сливаться друг с другом, делиться. Форма и размеры митохондрий животных клеток разнообразны, но в среднем толщина их около 0,5 мкм, а длина - от 1 до 10 мкм. Количество их в клетках сильно варьирует - от единичных элементов до сотен. Так, в клетке печени они составляют более 20% общего объема цитоплазмы. Площадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности её плазматической мембраны.
Митохондрии ограничены двумя мембранами толщиной около 7 нм. Наружная митохондриальная мембрана ограничивает собственно внутреннее содержимое митохондрии, её матрикс. Характерной чертой внутренних мембран митохондрий является их способность образовывать многочисленные впячивания внутрь митохондрий. Такие впячивания чаще имеют вид плоских гребней, или крист. Нити матрикса митохондрий представляют собой молекулы ДНК, а мелкие гранулы - митохондриальные рибосомы.
Эндоплазматическая сеть была открыта К.Р. Портером в 1945 г. Эта органелла представляет собой совокупность вакуолей, плоских мембранных мешков или трубчатых образований, создающих как бы мембранную сеть внутри цитоплазмы. Различают два типа - гранулярную и гладкую эндоплазматическую сеть.
Гранулярная эндоплазматическая сеть представлена замкнутыми мембранами, отличительной чертой которых является то, что они со стороны гиалоплазмы покрыты рибосомами. Рибосомы участвуют в синтезе белков, выводимых из данной клетки. Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков-ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения.
Белки, накапливающиеся в полостях сети, могут, минуя гиалоплазму, транспортироваться в вакуоли комплекса Гольджи, где они часто модифицируются и входят в состав либо лизосом, либо секреторных гранул.
Роль гранулярной эндоплазматической сети заключается в синтезе на её полисомах экспортируемых белков, в их изоляции от содержимого гиалоплазмы внутри мембранных полостей, в транспорте этих белков в другие участки клетки, а также в синтезе структурных компонентов клеточных мембран.
Агранулярная (гладкая) эндоплазматическая сеть также представлена мембранами, образующими мелкие вакуоли и трубки, канальцы, которые могут ветвиться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев обычно около 50-100 нм.
Гладкая эндоплазматическая сеть возникает и развивается за счет гранулярной эндоплазматической сети.
Деятельность гладкой ЭПС связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая ЭПС участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих стероиды в корковом веществе надпочечников и сустентоцитах ( клетки Сертоли) семенников.
В поперечнополосатых мышечных волокнах гладкая ЭПС способна депонировать ионы кальция, необходимые для функции мышечной ткани.
Очень важна роль гладкой ЭПС в дезактивации различных вредных для организма веществ.
Комплекс Гольджи (КГ). В 1898 г. К. Гольджи, используя свойства связывания тяжелых металлов с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал внутренним сетчатым аппаратом.
Он представлен мембранными структурами, собранными вместе в небольшой зоне. Отдельная зона скопления этих мембран называется диктиосомой. Таких зон в клетке может быть несколько. В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены 5-10 плоских цистерн, между которыми располагаются тонкие прослойки гиалоплазмы. Кроме цистерн в зоне КГ наблюдается множество мелких пузырьков (везикул). КГ участвует в сегрегации и накоплении продуктов, синтезированных в цитоплазматической сети, в их химических перестройках, созревании; в цистернах КГ происходит синтез полисахаридов, их комплексирование с белками и, главное, выведение готовых секретов за пределы клетки.
Лизосомы - это разнообразный класс шаровидных структур размером 0,2-0,4 мкм, ограниченных одиночной мембраной.
Характерным признаком лизосом является наличие в них гидролитических ферментов, расщепляющих различные биополимеры. Лизосомы были открыты в 1949 г. де Дьювом.
Пероксисомы - небольшие размером 0,3-1,5 мкм овальной формы тельца, ограниченные мембраной. Они особенно характерны для клеток печени, почек. Ферменты окисления аминокислот образуют перекись водорода, который разрушается ферментом каталаза. Каталаза пероксисом играет важную защитную роль, так как Н2О2 является токсическим веществом для клетки.
Немембранные органеллы
Рибосомы - элементарные аппараты синтеза белковых, полипептидных молекул - обнаруживаются во всех клетках. Рибосомы - это сложные рибонуклеопротеиды, в состав которых входят белки и молекулы РНК. Размер функционирующей рибосомы эукариотических клеток 25 х 20 х 20 нм.
Различают единичные рибосомы и комплексные рибосомы (полисомы). Рибосомы могут располагаться свободно в гиалоплазме и быть связанными с мембранами эндоплазматической сети. Свободные рибосомы образуют белки в основном на собственные нужды клетки, связанные обеспечивают синтез белков “” на экспорт”.
Микротрубочки относятся к фибриллярным компонентам белковой природы. В цитоплазме они могут образовывать временные образования (веретено деления). Микротрубочки входят в состав центриолей, а также являются основными структурными элементами ресничек и жгутиков. Они представляют собой прямые, неветвящиеся длинные полые цилиндры. Их внешний диаметр составляет около 24 нм, внутренний просвет - 15 нм, толщина сетки - 5 нм. Микротрубочки содержат белки - тубулины. Создавая внутриклеточный скелет, микротрубочки могут быть факторами ориентированного движения клетки в целом и её внутриклеточных компонентов, создавая факторы направленных потоков разных веществ.
Центриоли. Термин был предложен Т. Бовери в 1895 г. для обозначения очень мелких телец. Центриоли обычно расположенные в паре - диплосома, окружены зоной более светлой цитоплазмы, от которой отходят радиально тонкие фибриллы (центросфера). Совокупность центриолей и центросферы называют клеточным центром. Эти органеллы в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся клетках располагаются вблизи КГ.
Основой строения центриолей являются расположенные по окружности 9 триплетов микротрубочек, образующих таким образом полый цилиндр. Его ширина около 0,2 мкм, а длина 0,3-0,5 мкм.
Кроме микротрубочек в состав центриоли входят дополнительные структуры - “”ручки”, соединяющие триплеты. Системы микротрубочек центриоли можно описать формулой: (9 х 3) + 0, подчеркивая отсутствие микротрубочек в её центральной части.
При подготовке клеток к митотическому делению происходит удвоение центриолей.
Полагают, что центриоли участвуют в индукции полимеризации тубулином при образовании микротрубочек. Перед митозом центриоль является одним из центров полимеризации микротрубочек веретена клеточного деления.
Реснички и жгутики. Это специальные органеллы движения. В основании ресничек и жгутика в цитоплазме видны мелкие гранулы - базальные тельца. Длина ресничек 5-10 мкм, жгутиков - до 150 мкм.
Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с диаметром 200 нм. Он покрыт плазматической мембраной. Внутри расположена аксонема (“осевая нить”), состоящая из микротрубочек.
Аксонема в своём составе имеет 9 дуплетов микротрубочек. Здесь систему микротрубочек реснички опмсывают (9 х 2) + 2.
Свободные клетки, имеющие реснички и жгутики, обладают способностью двигаться. Способ их движения “”скользящие нити”.
К фибриллярным компонентам цитоплазмы относятся микрофиламенты толщиной 5-7 нм и так называемые промежуточные филаменты, микрофибриллы, толщиной около 10 нм.
Микрофиламенты встречаются во всех типах клеток. По строению и функциям они бывают разные, но отличить их морфологически друг от друга трудно. Химический состав их разный. Они могут выполнять функции цитоскелета и участвовать в обеспечении движения внутри клетки.
Промежуточные филаменты тоже белковые структуры. В эпителии в их состав входит кератин. Пучки филаментов образуют тонофибриллы, которые подходят к десмосомам. Роль промежуточных микрофиламентов скорее всего опорно-каркасная.
Включения цитоплазмы. Это необязательные компоненты клетки, возникающие и исчезающие в зависимости от метаболического состояния клеток. Различают включения трофические, секреторные, экскреторные и пигментные. Трофические включения - это нейтральные жиры и гликоген. Пигментные включения могут быть экзогенные (каротин, красители, пылевые частицы и др.) и эндогенные (гемоглобин, меланин и др.). Наличие их в цитоплазме может изменять цвет ткани. Нередко пигментация ткани служит диагностическим признаком.
Ядро обеспечивает две группы общих функций: одну, связанную собственно с хранением и передачей генетической информации, другую - с её реализацией, с обеспечением синтеза белка.
В ядре происходит воспроизведение или редупликация молекул ДНК, что дает возможность при митозе двум дочерним клеткам получить совершенно одинаковые в качественном и количественном отношении объемы генетической информации.
Другой группой клеточных процессов, обеспечиваемых активностью ядра, является создание собственного аппарата белкового синтеза. Это не только синтез, транскрипция на молекулах ДНК разных информационных РНК, но и транскрипция всех видов транспортных и рибосомальных РНК.
Таким образом, ядро является не только вместилищем генетического материала, но и местом, где этот материал функционирует и воспроизводиться.
Ядро неделящейся, интерфазной клетки обычно одно на клетку. Ядро состоит из хроматина, ядрышка, кариоплазмы (нуклеоплазмы) и ядерной оболочки, отделяющей его от цитоплазмы (кариолеммы).
Кариоплазма или ядерный сок - микроскопически бесструктурное вещество ядра. Он содержит различные белки (нуклеопротеиды, гликопротеиды), ферменты и соединения, участвующие в процессах синтеза нуклеиновых кислот, белков и др. веществ, входящих в состав кариоплазмы. Электронно - микроскопически в ядерном соке выявляют рибонуклеопротеидные гранулы 15 нм в диаметре.
В ядерном соке выявлены также гликолитические ферменты, участвующие в синтезе и расщеплении свободных нуклеотидов и их компонентов, энзимы белкового и аминокислотного обмена. Сложные процессы жизнедеятельности ядра обеспечиваются энергией, освобождающейся в процессе гликолиза, ферменты которого содержатся в ядерном соке.
Хроматин. В состав хроматина входит ДНК в комплексе с белком. Такими же свойствами обладают и хромосомы, которые отчетливо видны во время митотического деления клеток. Хроматин интерфазных ядер представляет собой хромосомы, которые теряют в это время свою компактную форму, разрыхляются, деконденсируются. Зоны полной деконденсации называются эухроматином; неполного разрыхления хромосом - гетерохроматином. Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде плотных хромосом.
Ядрышко. Это одно или несколько округлой формы тельца величиной 1-5 мкм, сильно преломляющих свет. Его ещё называют нуклеолой. Ядрышко - самая плотная структура ядра - является производным хромосомы.
В настоящее время известно, что ядрышко - это место образования рибосомальных РНК и полипептидных цепей в цитоплазме.
Ядрышко неоднородно по своему строению: в световом микроскопе можно видеть его тонковолокнистую организацию. В электронном микроскопе выделяют два основных компонента: гранулярный и фибриллярный. Фибриллярный компонент - это рибонуклеопротеидные тяжи предшественников рибосом, гранулы - созревающие субъединицы рибосом.
Ядерная оболочка состоит из внешней ядерной мембраны и внутренней мембраны оболочки, разделенных перинуклеарным пространством. Ядерная оболочка содержит ядерные поры. Мембраны ядерной оболочки в морфологическом отношении не отличаются от остальных внутриклеточных мембран.
Поры имеют диаметр около 80-90 нм. Поперёк поры имеется диафрагма. Размеры пор у данной клетки обычно стабильны. Число пор зависит от метаболической активности клеток: чем интенсивнее синтетические процессы в клетках, тем больше пор на единицу поверхности клеточного ядра.
Хромосомы. Как интерфазные, так и митотические хромосомы состоят из элементарных хромосомных фибрилл - молекул ДНК.
Морфологию митотических хромосом лучше всего изучать в момент их наибольшей конденсации, в метафазе и в начале анафазы. Хромосомы в этом состоянии представляют собой палочковидные структуры разной длины и довольно постоянной толщины. У большинства хромосом удается легко найти зону первичной перетяжки (центромеры), которая делит хромосому на два плеча. Хромосомы с равными или почти равными плечами называют метацентрическими, с плечами неодинаковой длины - субметацентрическими. Палочковидные хромосомы с очень коротким, почти незаметным вторым плечом называют акроцентрическими. В области первичной перетяжки расположен кинетохор. От этой зоны во время митоза отходят микротрубочки клеточного веретена. Некоторые хромосомы имеют, кроме того, вторичные перетяжки, располагающиеся вблизи одного из концов хромосомы и отделяющие маленький участок - спутник хромосом. В этих местах локализована ДНК, ответственная за синтез рибосомальных РНК.
Совокупность числа, размеров и особенностей строения хромосом называют кариотипом данного вида. Кариотип крупного рогатого скота - 60, лошади - 66, свиньи - 40, овцы - 54, человека - 46.
Время существования клетки как таковой, от деления до деления или от деления до смерти называют клеточным циклом (рис.2).
Весь клеточный цикл состоит из 4 отрезков времени: собственно митоза, предсинтетического, синтетического и постсинтетического периодов интерфазы. В период G1 начинается рост клеток за счет накопления клеточных белков, что определяется увеличением количества РНК на клетку. В S - периоде происходит удвоение количества ДНК на ядро и соответственно удваивается число хромосом. Здесь уровень синтеза РНК возрастает соответственно увеличению количества ДНК, достигая своего максимума в G2-периоде. В G2-периоде происходит синтез информационной РНК, необходимой для прохождения митоза. Среди синтезирующихся в это время белков особое место занимают тубулины - белки митотического веретена.
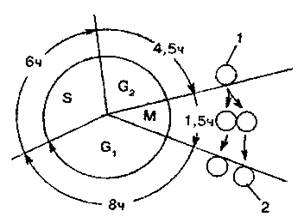
Рис. 2. Жизненный цикл клетки:
М – митоз; G1 - предсинтетический период; S — синтетический период; G2 — постсинтетический период; 1 - старая клетка (2n4c); 2- молодые клетки (2n2c)
Преемственность хромосомного набора обеспечивается клеточным делением, которое называется митозом. Во время этого процесса происходит полная перестройка ядра. Митоз состоит из последовательного ряда стадий, сменяющихся в определенном порядке: профазы, метафазы, анафазы и телофазы. В процессе митоза ядро соматической клетки делится таким образом, что каждая из двух дочерних клеток получает точно такой же набор хромосом, какой имела материнская.
Способность клеток к воспроизведению - важнейшее свойство живой материи. Благодаря этой способности обеспечивается непрерывная преемственность клеточных поколений, сохранение клеточной организации в эволюции живого, совершается рост и регенерация.