Біологія індивідуального розвитку
Міністерство Науки і Освіти України
ДНУ
ім. О. Гончара
Контрольна робота
З курсу «Біологія індивідуального розвитку»
Виконала
Студентка 3 курсу
Спеціальніть «Біологія»
Будко Г.О.
Дпіпропетровськ
2011р.
План
1. Способи гаструляції
2. Провізорні органи зародка у птахів
гаструляція провізорний зародок птах
1. Способи гаструляції
Характер гаструляції в тваринному царстві надзвичайно сильно варіює, проте здійснюється вона за участю порівняно нечисленних механізмів. При гаструляції спостерігається поєднання декількох основних типів рухів.
- інвагінація
Є укручуванням ділянки клітинної стінки зародка подібно до того, як вдавлюється всередину м'яка стінка гумового м'яча, коли на нього натискають. При інвагінації механічна цілісність стінки бластули не порушується. Впячивание однієї з ділянок бластодермы всередину здійснюється цілим пластом (мал. 1).
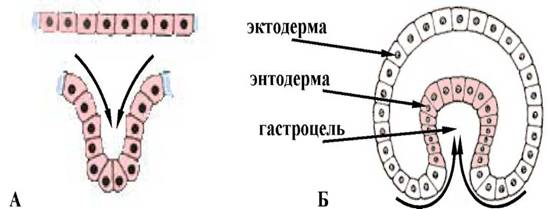
Мал. 1. Схема процесу інвагінації: А - рух пласта клітин; Б - инвагінаційна гаструла.
У ланцетника впячиваються клітини вегетативного полюса, а у земноводних інвагінація відбувається на межі між анимальным і вегетативним полюсом в області сірого серпа. Процес інвагінації можливий тільки в яйцях з невеликою або середньою кількістю жовтка. В результаті утворюється двуслойный мішок, зовнішньою стінкою якого є первинна ектодерма, а внутрішньою - первинна ентодерма. Бластоцель (первинна порожнина тіла або порожнина Бера) поступово витісняється, в ході впячивання утворюється гастроцель - первинний кишковик або архентерон, а отвір за допомогою якого він сполучається із зовнішнім середовищем, називають бластопором або первинним ротом. Останній з усіх боків оточений губами бластопора.
Доля бластопора у різних тварин неоднакова. У багатьох тварин первинний рот, розвиваючись і диференціюючись, перетворюється на рот дефінітива дорослого організму - первиннороті (черв'яки, молюски і членистоногі). Не менш велика група вторічноротих, у яких бластопор перетворюється на анальний отвір (щетинкочелюстні, плеченогие, голкошкірі, кишководишні) або в нервово-кишковий канал, що знаходиться в задньому кінці ембріона (хордові).
- інволюція
При цьому відбувається укручування всередину зародка зовнішнього пласта клітин, що збільшується в розмірах, який поширюється по внутрішній поверхні клітин (мал. 2), що залишаються зовні.
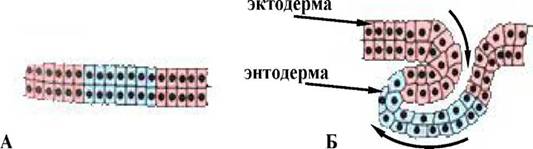
Мал. 2. Схема інволюції : А - початковий пласт клітин, Б - рухи клітинного пласта в ході інволюції
- імміграція
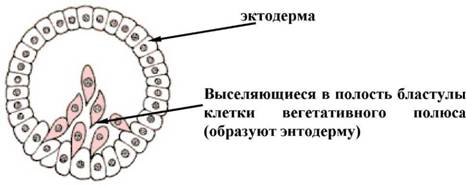
Є найбільш древній спосіб гаструляции, відкритий И.И. Мечниковим (1886). Імміграційна гаструла характерна для багатьох кишковополосних. Вона розвивається внаслідок активного виселення (ингрессии) частини клітин стінки бластули всередину бластоцеля. Це виселення клітин може відбуватися як з одного (вегетативного) полюса - уніполярна імміграція, так і з двох протилежних (анимального і вегетативного) полюсів - біполярна імміграція (мал. 3).
 _
_
Мал. 3. Уніполярна імміграція (схема)
Іноді процеси імміграції йдуть без особливого порядку по усій поверхні бластули - мультиполярная імміграція. Клітини, що виселилися, пізніше утворюють внутрішній шар гаструли - ентодерму. Таким чином, як і в ході інвагінації зародок стає двошаровим. Уніполярна імміграція характерна майже для усіх гідромедуз, особливо тих, яким властива плаваюча целобластула і мультиполярная імміграція, навпаки, зустрічаються набагато рідше. У багатьох кишковополостних, яким властива імміграційна гаструла, відбувається таке масове, активне виселення клітин бластодермы, що бластоцель повністю заповнюється їх щільною масою і зникає. Важливою особливістю імміграційної гаструли є відсутність бластопора, а значить немає і характерного для инвагінацонной гаструли сполучення гастроцеля із зовнішнім середовищем.
При імміграції відбувається переміщення окремих клітин або груп клітин, не об'єднаних в єдиний пласт. Імміграція, в тому або іншому виді, зустрічається у усіх зародків, але найбільшою мірою характерна для другої фази гаструляции вищих позвонкових (птахів і ссавців).
- делямінація
У тих випадках, коли дроблення закінчується утворенням бластули з невираженою або майже відсутньою порожниною (бластоцелем), наприклад морули у деяких кишковополостних, спостерігається розщеплення єдиного клітинного пласта на два більш менш паралельних. Кожна клітина, що утворює бластодерму, шляхом мітотичного ділення розщепляється і відшнуровує другий шар (мал. 4). гаструляції обмежується вирівнюванням внутрішніх стінок клітин зовнішнього шару, причому таке вирівнювання нерідко йде хвилею від однієї сусідньої клітини до іншої. Уздовж вирівняних поверхонь формується базальна мембрана, що відділяє зовнішній клітинний шар (ектодерму) від внутрішньої маси клітин, яка уся стає ентодермою. Делямінацію можна спостерігати у зародків з неповним типом дроблення, таких як плазуни птахи, нижчі ссавці.
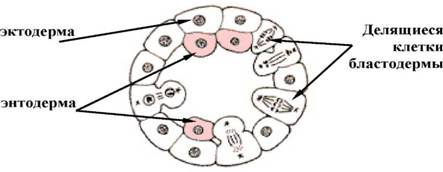
Мал. 4. Деляіинація (схема)
– епіболія
Представляє рух епітеліальних пластів клітин (зазвичай ектодермальних), які поширюються як одне ціле, а не індивідуально і оточують глибокі шари зародка. Іншими словами, відбувається обростання дрібними клітинами анимального полюса більш великих, відстаючих в швидкості ділення і менш рухливих, клітин вегетативного полюса (мал. 5).
Формування епіболічної гаструли яскраво виражене у земноводних і деяких організмів з різко телоцитальною будовою яєць. В результаті у зародків таких тварин бластопор відсутній і архентерон не формується. Тільки далі, коли макрозаходи ділячись стають меншого розміру, утворюється порожнина -формируется зачаток первинного кишковика.
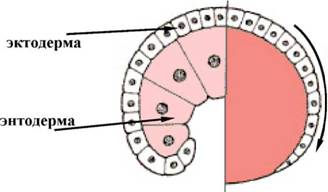
Мал. 5. Епіболія (схема)
У чистому вигляді вказані способи гаструляції зустрічаються украй рідко. У кожному конкретному випадку ембріогенезу, як правило, поєднуються декілька типів рухів. В результаті прийнято виділяти останній, змішаний, спосіб гаструляції.
2. Провізорні органи зародка у птахів
Провізорні, або тимчасові, органи утворюються в ембріогенезі ряду представників хребетних для забезпечення життєво важливих функцій, таких, як дихання, живлення, виділення, рух та ін. Недорозвинені органи самого зародка ще не здатні функціонувати за призначенням, хоча обов'язково грають якусь роль в системі цілісного організму, що розвивається. Як тільки зародок досягає необхідної міри зрілості, коли більшість органів здатні виконувати життєво важливі функції, тимчасові органи розсмоктуються або відкидаються.
Час утворення провізорних органів залежить від того, які запаси поживних речовин були накопичені в яйцеклітині і в яких умовах середовища відбувається розвиток зародка. У безхвостих земноводних, наприклад, завдяки достатній кількості жовтка в яйцеклітині і тому, що розвиток йде у воді, зародок здійснює газообмін і виділяє продукти дисиміляції безпосередньо через оболонки яйця і досягає стадії пуголовка. На цій стадії утворюються провізорні органи дихання (зябра), травлення і рухи, пристосовані до водного способу життя. Перераховані личинкові органи дають можливість пуголовку продовжити розвиток. Після досягнення стану морфофункциональной зрілості органів дорослого типу тимчасові органи зникають в процесі метаморфоза.
У плазунів і птахів запасів жовтка в яйцеклітині більше, але розвиток йде не у воді, а на суші. У зв'язку з цим дуже рано виникає потреба в забезпеченні дихання і виділення, а також в захисті від висихання. У них вже в ранньому ембріогенезі, майже паралельно з нейруляцією, починається формування провізорних органів, таких, як амніон, хоріон і жовтковий мішок. Трохи пізніше формується аллантоис. У ссавців плацентарних ці ж провізорні органи утворюються ще раніше, оскільки в яйцеклітині дуже мало жовтка. Розвиток таких тварин відбувається внутріутробний, утворення провізорних органів у них співпадає за часом з періодом гаструляції.
Наявність або відсутність амніона і інших провізорних органів лежить в основі ділення хребетних на дві групи: Amniota і Anamnia. Еволюційно більш древні хребетні, такі, що розвиваються виключно у водному середовищі і представлені такими класами, як Круглоротые, Риби і Земноводні, не потребують додаткових водних і інших оболонках зародка і складають групу анамний. До групи амниот відносять первичноназемных хребетних, тобто тих, у кого ембріональний розвиток протікає в наземних умовах.
Це три класи: Плазуни, Птахи і Ссавці. Вони є вищими хребетними, оскільки мають скоординированные і високоефективні системи органів, що забезпечують їм існування в найбільш складних умовах, якими є умови суші. Ці класи налічують велику кількість видів, що повторно перейшли у водне середовище. Таким чином, вищі хребетні виявилися в змозі освоїти усі місця існування. Подібна досконалість була б неможливою, у тому числі і без внутрішнього запліднення і спеціальних провізорних ембріональних органів.
У будові і функціях провізорних органів різних амниот багато спільного. Характеризуючи в найзагальнішому вигляді провізорні органи зародків вищих хребетних, звані також зародковими оболонками, слід зазначити, що усі вони розвиваються з клітинного матеріалу зародкових листків, що вже сформувалися. Деякі особливості є в розвитку зародкових оболонок ссавців плацентарних, про що буде сказано нижче.
Амніон є ектодермальним мішком, що укладає зародок і заповнений амниотической рідиною. Амниотическая оболонка спеціалізована для секреції і поглинання амниотической рідини, що омиває зародок. Амніон грає первинну роль в захисті зародка від висихання і від механічних ушкоджень, створюючи для нього найбільш сприятливе і природне водне середовище. Амніон має і мезодермальный шар з позазародкової соматоплеври, який дає початок гладким м'язовим волокнам. Скорочення цих м'язів викликають пульсацію амніона, а повільні коливальні рухи, що повідомляються при цьому зародку, мабуть, сприяють тому, що його зростаючі частини не заважають один одному.
Хоріон (сероза) - сама зовнішня зародкова оболонка, прилегла до шкаралупи або материнських тканин, виникаюча, як і амніон, з ектодерми і соматоплеври. Хоріон служить для обміну між зародком і довкіллям. У яйцеродних видів основна його функція - дихальний газообмін; у ссавців він виконує набагато більші функції, беручи участь окрім дихання в живленні, виділенні, фільтрації і синтезі речовин, наприклад гормонів.
Жовтковий мішок має энтодермальное походження, покритий вісцелярною мезодермою і безпосередньо пов'язаний з кишковою трубкою зародка. У зародків з великою кількістю жовтка він бере участь в живленні. У птахів, наприклад в спланхноплевре жовткового мішка, розвивається судинна мережа. Жовток не проходить через жовткову протоку, що сполучає мішок з кишкою. Спочатку він переводиться в розчинну форму під дією травних ферментів, що продукуються энтодермальными клітинами стінки мішка. Потім потрапляє в посудини і з кров'ю розноситься по усьому тілу зародка.
Доля жовткового мішка у різних тварин дещо різна. У птахів до кінця періоду інкубації залишки жовткового мішка вже знаходяться усередині зародка, після чого він швидко зникає і до кінця 6-ої доби після вилуплення повністю розсмоктується.
Аллантоис розвивається дещо пізніше за інші позазародкові органи. Він є мішковидним вирощуванням вентральної стінки задньої кишки. Отже, він утворений ентодермою зсередини і спланхноплеврой зовні. У рептилій і птахів аллантоис швидко доростає до хоріону і виконує декілька функцій. Передусім це вмістище для сечовини і сечової кислоти, які є кінцевими продуктами обміну азотвмісних органічних речовин. У аллантоисе добре розвинена судинна мережа, завдяки чому разом з хоріоном він бере участь в газообміні. При вилупленні зовнішня частина аллантоиса відкидається, а внутрішня - зберігається у вигляді сечового міхура.
Доступніше і простіше вивчити утворення і будову зародкових оболонок на прикладі зародка курки. На стадії нейрули три зародкові листки безпосередньо переходять від зародка до позазародкової частини, ніяк не відмежовуючись. У міру того як зародок набуває форми, навколо нього утворюється декілька складок, які як би підсікають зародок, відділяють його від жовтка і встановлюють чіткі межі між зародком і позазародковими областями. Вони називаються тулубовими складками. (Мал.1).
Спочатку утворюється головна складка. Вона підсікає знизу головну частину. Задні кінці цієї складки переходжуватимуть в бічні тулубові складки, що відмежовують тулуб зародка з боків. Хвостова складка відмежовує задній кінець зародка. Поступово звужується ніжка, що сполучає середню кишку і жовтковий мішок, утворюється передній і задній відділи кишки.
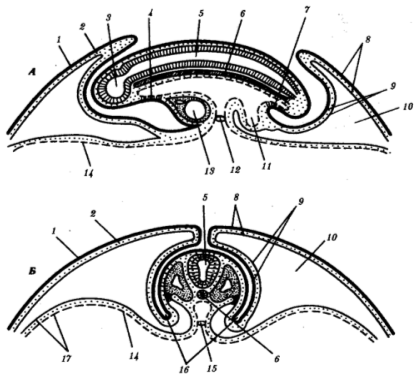
Мал. 1 Утворення тулубових складок і зародкових оболонок у зародка курчати. А -продольный зріз; Би -поперечный зріз:
1 -эктодерма, 2 -мезодерма, 3 -зачаток мозку, 4 -глоточная мембрана, 5 -нервная трубка, 6-хорда, 7-клоачна мембрана, 8-хорион, 9-амнион, 10-экзоделом, 11-аллаитоис, 12-область пупка, 13-зачаток серця, 14-энтодерма, 15-закладка кишковика, 16-тулубові складки, 17-жовтковий мішок
Одночасно з ектодерми і прилеглої до неї соматоплеври утворюється спочатку головна складка (мал.), яка, як капюшон, наростає на зародок згори. Кінці головної складки утворюють з боків амниотические валики. Вони ростуть поверх зародка один назустріч одному і зростаються, утворюючи відразу стінки амніона, прилеглого до зародка, і хоріону, що лежить зовні.
Пізніше утворюється аллантоис (мал.2). Загальний вигляд курячого зародка на 6-у добу інкубації зображений на мал.2. У різних ссавців процеси утворення провізорних органів більше або менше схожі на описаних вище у птахів.
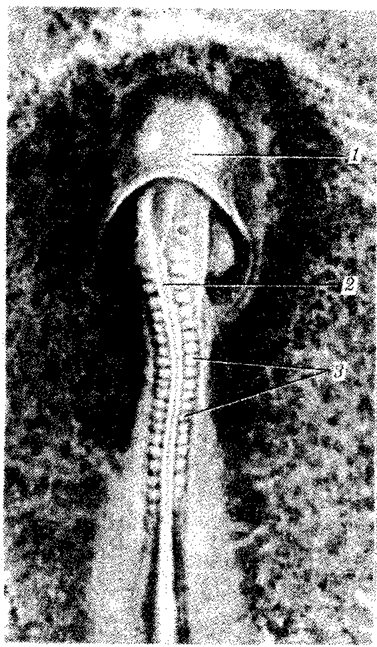
Мал. 2 Курячий зародок близько 40 ч інкубації: 1-головна складка амніона, 2-нервова трубка, 3 - соміти
Список літератури
1. Грин Н., Стаут У., Тейлор Д. Біологія / Н. Грин, У. Стаут, Д. Тейлор, під ред. Р. Сопера. - М.: Світ, 2007. - Т. 1. - 368 с.
2. Грин Н., Стаут У., Тейлор Д. Біологія / Н. Грин, У. Стаут, Д. Тейлор, під ред. Р. Сопера. - М.: Світ, 2009. - Т. 2. - 325 с.
3. М.А. Андрейчин, В.С. Копча Епідеміологія Терн. "Укр.медкнига"-2000р. стор. 349-356.
4. MED/96217734. Jacobsen H, Hanggi M, Ott M, Duncan IB, Owen S, Andreoni M, Vella S, Mous J. In vivo resistance to a human immunodeficiency virus type 1 proteinase inhibitor: mutations, kinetics, and frequencies. J Infect Dis. 1996 Jun;173(6):1379-87.