Филогенез и онтогенез коры больших полушарий
Общая информация о коре больших полушарий
Строение коры
Новая кора
Ассоциативные зоны и локализация полей
Филогенез коры
Онтогенез коры
Формирование коры в ходе внутриутробного развития ребенка
Первичные, вторичные и третичные поля коры
Заключение
Библиография
филогенез онтогенез кора полушарие
Введение
Кора головного мозга осуществляет тончайшее равновесие между организмом и внешней средой, регулирует и направляет физиологические процессы, протекающие внутри организма, обеспечивает его сложнейшее функциональное единство. В рамках данного реферата ставится цель посмотреть на процессы развития коры головного мозга в рамках большого периода филогенетического развития коры, а также индивидуального развития коры в рамках онтогенеза. Также ставится цель сравнить данные процессы с целью нахождения общих принципов и сходных процессов.
Общая информация о коре больших полушарий
Большие полушария – парные образования головного мозга. У человека они достигают ≈ 80% от общей массы мозга. Большие полушария осуществляют регуляцию высших нервных функций, лежащих в основе всех психических процессов человека, в то время как стволовая часть мозга обеспечивает низшие функции нервной системы, связанные с регуляцией деятельности внутренних органов.
Высшие функции обеспечиваются деятельностью особого отдела больших полушарий – коры головного мозга, которая несет главную ответственность за формирование условно-рефлекторных реакций. У человека по сравнению с животными кора одновременно ответственна и за согласование работы внутренних органов. Такое возрастание роли коры в регуляции всех функций в организме называется кортикализацией функций.
Кора выполняет следующие функции:
1. взаимодействие организма с внешней средой за счет безусловных и условных рефлексов.
2. осуществление высшей нервной деятельности (поведения) организма.
3. выполнение высших психических функций (мышления и сознания).
4. регуляция работы внутренних органов и обмена веществ в организме.
Кора больших полушарий представлена 12-18 миллиардами клеток, расположенных тонким слоем 3-4 мм на площади 2400 см2. 65-70 % этой площади находится в глубине борозд, а 30-35 % - на видимой поверхности полушарий. Кора состоит из нервных клеток, их отростков и нейроглинов, для которых характерно обилие межнейронных связей.
Строение коры
Кора покрывает поверхность больших полушарий с ее многочисленными бороздами и извилинами, за счет которых площадь коры значительно увеличивается. Различают ассоциативные зоны коры, а также сенсорную и моторную кору – области, в которых сосредоточены нейтроны, иннервирующие различные части тела.
Кора больших полушарий связана нервными путями со всеми нижележащими отделами центральной нервной системы, а через них — со всеми органами тела. С одной стороны, импульсы, поступающие с периферии, доходят до той или иной точки коры, с другой — кора посылает «распоряжения» в нижележащие отделы мозга, а оттуда — к различным органам.
По происхождению и структуре кора больших полушарий неоднородна. Большую часть коры у человека занимает новая кора неокортекс (neocortex), филогенетически наиболее молодая корковая формация. Филогенетически более ранние корковые структуры — древняя кора (paleocortex) и старая кора (archicortex) — занимают небольшую часть поверхности полушарий. Закладка новой коры образуется в латеральных частях плаща. Новая кора интенсивно развивается и оттесняет древнюю кору на основание полушарий, где она сохраняется в виде узкой полоски обонятельной коры и занимает 0,6% поверхности коры на вентральной поверхности полушарий, а старая кора отодвигается на медиальные поверхности полушарий, занимает 2, 2% поверхности коры и представлена гиппокампом и зубчатой извилиной. По происхождению и клеточному строению новая кора отличается от древней и старой коры. Переход от одной корковой формации к другой в клеточном строении происходит постепенно. Кора переходного типа называется межуточной корой, она занимает 1, 3% общей площади коры. Таким образом, большую часть поверхности коры (95, 6%) занимает новая кора.
Для древней коры характерно отсутствие послойного строения. В ней преобладают крупные нейроны, сгруппированные в клеточные островки. Старая кора имеет три клеточных слоя. Ключевой структурой старой коры является гиппокамп. Гиппокамп имеет обширные связи со многими другими структурами мозга. Он является центральной структурой лимбической системы мозга.
Функциональной единицей коры является вертикальная колонка взаимосвязанных нейронов. Все нейроны вертикальной колонки отвечают на одно и тоже афферентное раздражение одинаковой реакцией и совместно формируют эфферентный ответ. Распространение возбуждения в горизонтальном направлении (иррадиация) обеспечивается поперечными волокнами, идущими от одной вертикальной колонки к другой, а ограничивается – процессами торможения. Возникновение возбуждения в вертикальной колонке нейронов приводит к активности спинальные мотонейроны и к сокращению связанных с ними мышц.
Упорядоченное положение клеток в коре называется цитоархитектоникой, а их волокон – миелоархитектоникой.
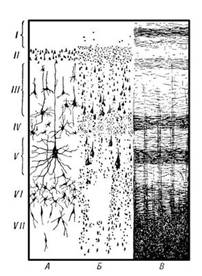
Рисунок 1. Схема цитоархитектонического строения новой коры головного мозга
На рисунке 1 представлена схема цитоархитектонического строения новой коры. В разделе А окрашены только тела клеток, Б окрашены отдельные клетки с отростками; В окрашены волокна. Слои коры: I — молекулярный слой; II — наружный зернистый слой; III — пирамидный слой; IV — внутренний зернистый слой; V — ганглионарный слой; VI — полиморфный слой.
Новая кораВсе области новой коры построены по единому принципу. Исходным типом является шестислойная кора (рис. 1). Слои представлены следующим образом:
Iслой — самый поверхностный, толщиной около 0,2 мм, называется молекулярным (lamina molecularis). Он состоит из волокон апикальных дендритов и аксонов, поднимающихся от клеток нижних слоев, которые контактируют друг с другом. Нейронов в молекулярном слое незначительное количество. Это мелкие горизонтальные клетки и клетки-зерна. Все отростки клеток молекулярного слоя располагаются в пределах этого же слоя.
IIслой — наружный зернистый (lamina granulans externa).Толщина наружного зернистого слоя — 0,10 мм. Он состоит из мелких пирамидных и звездчатых нейронов. Аксоны этих нейронов оканчиваются на нейронах III, V и VI слоев.
III слой — пирамидный {lamina pyramidalis),толщиной около 1 мм, состоит из мелких и средних пирамидных клеток. Типичный пирамидный нейрон имеет форму треугольника, вершина которого направлена вверх. От вершины отходит апикальный дендрит, ветвящийся в вышележащих слоях. Аксон пирамидной клетки отходит от основания клетки и направляется вниз. Дендриты клеток III слоя направляются во второй слой. Аксоны клеток III слоя оканчиваются на клетках нижележащих слоев или образуют ассоциативные волокна.
IV слой — внутренний зернистый {lamina granulans internus).Он состоит из звездчатых клеток, имеющих короткие отростки, и малых пирамид. Дендриты клеток IV слоя уходят в молекулярный слой коры, а их коллатерали ветвятся в своем слое. Аксоны клеток IV слоя могут подниматься в вышележащие слои или уходить в белое вещество как ассоциативные волокна. Толщина IV слоя от 0,12 до 0,3 мм.
V слой — ганглионарный {lamina ganglionaris) — слой больших пирамид. Самые крупные клетки коры расположены именно в этом слое (гигантские пирамиды Беца передней центральной извилины). Их апикальные дендриты достигают молекулярного слоя, а базальные дендриты распределяются в своем слое. Аксоны клеток V слоя покидают кору и являются ассоциативными, комиссуральными или проекционными волокнами. Толщина V слоя достигает 0,5 мм.
VI слой коры — полиморфный {lamina multiformis).Содержит клетки разнообразной формы и размера, имеет толщину от 0,1 до 0,9 мм. Часть дендритов клеток этого слоя достигает молекулярного слоя, другие же остаются в пределах IV и V слоев. Аксоны клеток VI слоя могут подниматься к верхним слоям или уходить из коры в качестве коротких или длинных ассоциативных волокон.
Клетки одного слоя коры выполняют сходную функцию в обработке информации. I и IV слои являются местом ветвления ассоциативных и комиссуральных волокон, т.е. получают информацию от других корковых структур. III и IV слои являются входными, афферентными для проекционных полей, так как именно в этих слоях заканчиваются таламические волокна. V слой клеток выполняет эфферентную функцию, его аксоны несут информацию к нижележащим структурам мозга. VI слой также является выходным, но его аксоны кору не покидают, а являются ассоциативными.
Основным принципом функциональной организации коры является объединение нейронов в колонки. Колонка расположена перпендикулярно поверхности коры и охватывает все ее слои от поверхности к белому веществу. Связи между клетками одной колонки осуществляются по вертикали вдоль оси колонки. Боковые отростки клеток имеют небольшую длину. Связь между колонками соседних зон осуществляется через волокна, уходящие вглубь, а затем входящие в другую зону, т.е. короткие ассоциативные волокна. Функциональная организация коры в виде колонок обнаружена в соматосенсорной, зрительной, моторной и ассоциативной коре. Отдельные зоны коры имеют принципиально одинаковое клеточное строение, однако есть и отличия, особенно в структуре III, IV и V слоев, которые могут распадаться на несколько подслоев. Кроме этого, существенными цитоархитектоническими признаками являются плотность расположения и размеры клеток, наличие специфических типов нейронов, расположение и направление хода миелиновых волокон.
Цитоархитектонические особенности позволили разделить всю поверхность коры на 11 цитоархитектонических областей, включающих в себя 52 поля (по Бродману). Каждое цитоархитектоническое поле обозначено на картах мозга номером, который присваивался ему в порядке описания.
В первом слое коры волокна образуют полоску молекулярной пластинки. Во втором слое залегают тонкие волокна наружной зернистой пластинки. В составе четвертого слоя коры находится полоска внутренней зернистой пластинки (наружная полоска Байярже). В пятом слое содержатся волокна внутренней пирамидной пластинки (внутреняя полоска Байярже).
Основная информация в кору поступает по специфическим афферентным проводящим путям, заканчивающимся на клетках 3 и 4 слоев. Неспецифические пути от РФ заканчиваются в верхних слоях коры и регулируют ее функциональное состояние (возбуждение, торможение).
Звездчатые нейроны выполняют главным образом чувствительную (афферентную) функцию. Пирамидные и веретеновидные клетки – это преимущественно двигательные (эфферентные) нейроны.
Часть клеток коры принимают информацию от любых рецепторов организма – это полисенсорные нейроны, воспринимающие импульсы только от определенных рецепторов (зрительных, слуховых, тактильных и т.д.). Клетки нейроглии выполняют вспомогательные функции: трофическую, нейросекреторную, защитную, изолирующую.
Специализированные нейроны и другие клетки, входящие в состав вертикальных колонок, образуют отдельные участки коры, которые называются проекционными зонами (цитоархитектоническими полями)(1) . Эти функциональные зоны коры делятся на 3 группы:
– афферентные (чувственные);
– эфферентные (двигательные или моторные);
– ассоциативные (соединяют предыдущие зоны и обусловливают сложную работу мозга, лежащую в основе высшей психической деятельности).
Ассоциативные зоны и локализация полейКаждый анализатор (например, зрения, обоняния, слуха и т.д.) имеет, по представлению И. П. Павлова, в коре головного мозга центральную часть (ядро), где осуществляется высший анализ и синтез, и широкую периферическую зону, в которой аналитические и синтетические процессы совершаются в элементарном виде. Между ядрами отдельных анализаторов разбросаны и перемешаны нервные элементы, принадлежащие различным анализаторам. Если ядро анализатора в силу каких-либо причин разрушено или выбыло из строя, его функцию перенимают периферические элементы того же анализатора. Современная физиология отвергает и узкий («абсолютный») локализационизм и принцип однородности, равноценности всех участков коры мозга. Локализация существует, но имеет «подвижный», «динамический» характер, о чем еще много лет назад говорил И.П. Павлов.
Нервные образования, которые мы привыкли называть «центрами», не ограничиваются корой головного мозга. Они включают и подкорковые структуры, значение которых необычайно велико.
Следует помнить, что любой центр коры головного мозга теснейшим образом связан со всеми другими отделами центральной нервной системы. В этом объединении, или, как говорят, интеграции, и заключается ведущая роль коры мозга в организме. Представление о единых корковых центрах, полностью обеспечивающих какую-либо определенную функцию, является в настоящее время пройденным этапом в физиологии.
К тому же кора головного мозга отличается необычайной пластичностью, и одни отделы ее легко перенимают функции других, компенсируя расстройство их деятельности, вызванное различными причинами. Наиболее важная задача современной науки выявить анатомическую основу физиологических процессов и одновременно установить связи и взаимосвязи между всеми явлениями, наблюдаемыми в головном мозгу. Исследования, проведенные различными авторами, как отечественными, так и зарубежными, показали, что в центральной извилине мозга, расположенной спереди от центральной борозды, находится специальная двигательная область. Раздражение ее электрическим током вызывает сокращение определенных мышц противоположной стороны тела. Напротив, удаление этой области хирургическим путем ведет к расстройству координированных движений, шаткости походки, ослаблению мышц. У человека ранение двигательной области сопровождается обычно параличами и другими тяжелыми нарушениями деятельности организма. С помощью метода условных рефлексов удалось показать, что так называемые двигательные центры содержат чувствительные клетки, к которым приходят периферические раздражения от двигательного аппарата (костей, суставов, мышц). Эта область является мозговым концом двигательного анализатора в такой же степени, как затылочная — мозговым концом зрительного анализатора, височная — слухового анализатора и т.д. В двигательной области имеются как чувствительные клетки, расположенные в верхних слоях коры, так и двигательные, сосредоточенные в ее нижних слоях. Импульсы от рецепторов двигательного аппарата поступают в чувствительные клетки передней мозговой извилины, а отсюда уже передаются двигательным клеткам головного и спинного мозга.
У человека ассоциативные зоны достигают наибольшего развития. Локализация функций в коре головного мозга относительна – здесь нельзя провести каких-либо четких границ, поэтому мозг обладает высокой пластичностью, приспосабливаемостью к повреждениям. Тем не менее, морфологическая и функциональная неоднородность коры позволила выделить в ней 52 цитоархитектонических поля (К. Бродман), а среди них – центры зрения, слуха, осязания и др. Все они связаны между собой волокнами проводящих путей белого вещества, которые делятся на 3 типа:
1. ассоциативные (связывают зоны коры в пределах одного полушария),
2. комиссуральные (связывают симметричные зоны коры двух полушарий через мозолистое тело),
3. проекционные (связывают кору и подкорку с периферическими органами, бывают чувствительные и двигательные).
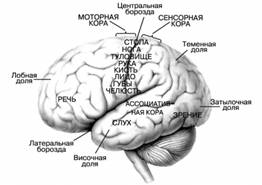
Рисунок 2. Значение важнейших зон коры головного мозга
1. Чувствительная зона коры (в постцентральной извилине) воспринимает импульсы от тактильных, температурных и болевых рецепторов кожи, а также от проприорецепторов противоположной половины тела.
2. Двигательная зона коры (в предцентральной извилине) содержит в 5 слое коры пирамидные клетки Беца, от которых идут импульсы произвольных движений к скелетным мышцам противоположной половины тела.
3. Премоторная зона (в основании средней лобной извилины) обеспечивает сочетанный поворот головы и глаз в противоположную сторону.
4. Праксическая зона (в надкраевой извилине) обеспечивает сложные целенаправленные движения практической деятельности и профессиональных двигательных навыков. Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
5. Центр проприоцептивного гнозиса (в верхней теменной дольке) обеспечивает восприятие импульсов проприорецепторов, контролирует ощущения тела и его частей как целостного образования.
6. Центр чтения (в верхней теменной дольке, вблизи затылочной доли) контролирует восприятие написанного текста.
7. Слуховая зона коры (в верхней височной извилине) воспринимает информацию от рецепторов органа слуха.
8. Слуховой центр речи, центр Вернике (в основании верхней височной извилины). Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
9. Слуховой центр пения (в верхней височной извилине). Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
10. Двигательный центр устной речи, центр Брока (в основании нижней лобной извилины) контролирует произвольные сокращения мышц, участвующих в речеобразовании. Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
11. Двигательный центр письменной речи (в основании средней лобной извилины) обеспечивает произвольные движения, связанные с написанием букв и других знаков. Зона асимметрична (у правшей – в левом, а у левшей – в правом полушарии).
12. Стереогностическая зона (в угловой извилине) контролирует узнавание предметов наощупь (стереогноз).
13. Зрительная зона коры (в затылочной доле) воспринимает информацию от рецепторов органа зрения.
14. Зрительный центр речи (в угловой извилине) контролирует движение губ и мимику говорящего оппонента, тесно связан с другими сенсорными и моторными речевыми центрами. Речь и сознание – это филогенетические наиболее молодые функции мозга, поэтому речевые центры имеют большое число рассеянных элементов и наименее локализованы. Речевые и мыслительные функции выполняются при участии всей коры. Речевые центры у человека сформировались на основе трудовой деятельности, поэтому они асимметричные, непарные и связаны с рабочей рукой.
Позади центральной борозды расположена чувствительная область коры. В ней заканчивается путь, начавшийся в рецепторах кожи и внутренних органов. Здесь расположен его конечный этап. Каждое полушарие мозга связано в основном с противоположной половиной тела. Однако существуют связи полушария и с одноименной половиной тела.
Филогенез корыКора — продукт длительного эволюционного развития, в процессе которого сначала появляется древняя кора, возникающая в связи с развитием обонятельного анализатора у рыб. С выходом животных из воды на сушу начинает интенсивно развиваться т. н. плащевидная, полностью обособленная от подкорки часть коры, которая состоит из старой и новой коры. Становление этих структур в процессе приспособления к сложным и разнообразным условиям наземного существования связано (совершенствованием и взаимодействием различных воспринимающих и двигательных систем. У земноводных кора представлена древней и зачатком старой коры, у пресмыкающихся хорошо развиты древняя и старая кора и появляется зачаток новой коры. Наибольшего развития новая кора достигает у млекопитающих, а среди них у приматов (обезьяны и человек), хоботных (слоны) и китообразных (дельфины, киты). В связи с неравномерностью роста отдельных структур новой коры её поверхность становится складчатой, покрываясь бороздами и извилинами. Совершенствование коры конечного мозга у млекопитающих неразрывно связано с эволюцией всех отделов центральной нервной системы. Этот процесс сопровождается интенсивным ростом прямых и обратных связей, соединяющих корковые и подкорковые структуры. Т. о., на более высоких этапах эволюции функции подкорковых образований начинают контролироваться корковыми структурами. Данное явление получило название кортиколизации функций. В результате кортиколизации ствол мозга образует с корковыми структурами единый комплекс, а повреждение коры на высших этапах эволюции приводит к нарушению жизненно важных функций организма. Наибольшие изменения и увеличение в процессе эволюции новой коры претерпевают ассоциативные зоны, в то время как первичные, сенсорные поля уменьшаются по относительной величине. Разрастание новой коры приводит к вытеснению старой и древней на нижнюю и срединную поверхности мозга.
У низших позвоночных (круглоротые и рыбы) развитие конечного мозга идет по пути утолщения основания, в котором формируются базальные ганглии в виде больших парных выпуклых масс. Боковые стенки и крыша конечного мозга, образующие плащ (мантию), остаются тонкими и повторяют форму полосатых тел в виде двух полушарий, хотя на два полушария конечный мозг еще не разделен, не разделена еще и полость его желудочка. В передней стенке конечного мозга развивается парное выпячивание — обонятельные доли. Они продолжаются в более или менее длинный стебелек, заканчивающийся обонятельной луковицей. Серое вещество мантии выстилает полость желудочка, а ее поверхность содержит белое вещество. В концевой пластинке располагается передняя комиссура, связывающая между собой полосатые тела. У двоякодышащих рыб и первых наземных позвоночных (амфибии) передняя стенка мозгового пузыря вдвигается посередине между полосатыми телами в виде продольной складки, разделяющей конечный мозг на два полушария. Полость мозгового пузыря разделяется на два боковых желудочка, которые соединяются сзади Монроевыми отверстиями с III желудочком. Мантия еще тонкая, но нервные клетки перемещаются к поверхности, однако остаются под слоем белого вещества. Над передней комиссурой появляется еще одна комиссура, соединяющая между собой мантийные структуры полушарий (аналог комиссуры гиппокампа). У рептилий конечный мозг разрастается еще больше, но это разрастание идет в основном за счет развития базальных ганглиев. Мантия остается тонкой, но на ее поверхности впервые появляется серое вещество, которое представляет собой кору. Эта кора является высшей обонятельной структурой и сравнима с гиппокампом млекопитающих. Усложнение организации конечного мозга млекопитающих происходит за счет развития мантии.
Мантия покрыта корой. Кроме древней и старой коры в филогенетическом ряду млекопитающих появляется новая кора. Чем более высокоразвит вид млекопитающих, тем большую поверхность занимают структуры новой коры. Вместе с развитием мантии преобразуются и полости желудочков. В добавление к имеющимся комиссурам появляется мозолистое тело, связывающее структуры новой коры обоих полушарий. С увеличением поверхности новой коры нарастает количество волокон мозолистого тела и его мощность. Мозолистое тело значительно разрастается и сдвигает назад комиссуру гиппокампа. Кора у низших млекопитающих развивается в связи с обонятельной сенсорной системой. У высших млекопитающих (обезьяны, человека) с обонянием связаны только структуры древней и старой коры. Кроме того, уже на ранних этапах развития позвоночных (амфибии и рептилии) кора получает афферентацию от сенсорных систем других модальностей через ядра таламуса. У млекопитающих кроме специфических ядер таламуса возникают и развиваются ассоциативные ядра, имеющие связи с неокортексом и определяющие развитие конечного мозга.
Третьим источником афферентных воздействий на кору больших полушарий является гипоталамус, который играет роль высшего регуляторного центра вегетативных функций. У млекопитающих филогенетически более древние отделы переднего гипоталамуса связаны с древней формацией коры и с древними ядрами амигдалы. Молодые отделы заднего гипоталамуса проецируются в старую кору.
Онтогенез корыКорковая пластинка появляется в процессе внутриутробного развития человека сравнительно рано — на 2-м месяце. Раньше всего выделяются нижние слои коры (VI—VII), затем — более высоко расположенные (V, IV, III и II; см. рис. 1). К 6 месяцам у эмбриона уже имеются все цитоархитектонические поля коры, свойственные взрослому человеку. После рождения в росте коры можно выделить три переломных этапа: на 2—3-м месяце жизни, в 2,5—3 года и в 7 лет. К последнему сроку цитоархитектоника коры полностью сформирована, хотя тела нейронов продолжают увеличиваться до 18 лет. Корковые зоны анализаторов завершают своё развитие раньше, и степень их увеличения меньше, чем у вторичных и третичных зон. Отмечается большое разнообразие в сроках созревания корковых структур у разных индивидуумов, что совпадает с разнообразием сроков созревания функциональных особенностей коры.
В начальной стадии формирования мозга человека передний мозг является закругленным концом нервной трубки. На этом этапе конечный мозг представлен тонкой ростральной стенкой переднего мозга. Затем эта стенка выпячивается в дорсолатеральном направлении и образует два мозговых пузыря. Полости этих пузырей образуют боковые желудочки. На ростральном конце мозговой трубки и III желудочка находится участок ткани, соединяющий оба пузыря конечного мозга. В дальнейшем он станет конечной пластинкой.
Дно мешка конечного мозга утолщается очень быстро, это утолщение образует полосатое тело (стадия 10-20 мм). В дальнейшем оно делится на хвостатое ядро, скорлупу, бледный шар и миндалину. По мере роста полушарий базальные ядра смещаются медиально и примерно на 10-й неделе развития (40 мм) сливаются с промежуточным мозгом.
Растущие аксоны покидают кору примерно на 8-й неделе развития (стадия зародыша — 23 мм). Волокна, которые не заканчиваются в полосатом теле, идут вдоль плоскости слияния основания конечного мозга с промежуточным, образуя внутреннюю капсулу. Волокна, идущие каудально и огибающие снизу средний мозг, образуют ножки мозга. Затем на вентральной поверхности продолговатого мозга они образуют пирамиды. Уходя в спинной мозг, пирамидные пути перекрещиваются и оканчиваются на мотонейронах спинного мозга.
Ассоциативные проводящие пути начинают выявляться в конце второго месяца развития. Свод появляется из гиппокампа в конце третьего месяца развития. Мозолистое тело появляется в начале четвертого месяца развития. Оно начинает формироваться в концевой пластинке в виде пучка поперечных волокон, лежащих над комиссурой гиппокампа. Мозолистое тело растет очень быстро в каудальном направлении и смещает туда же комиссуру гиппокампа и свод. Гиппокамп также смещается назад.
Пузыри конечного мозга разрастаются в ростральном, дорсальном и каудальном направлениях. На ранних стадиях развития (до 3—4-го месяца) пузыри имеют очень тонкую стенку и утолщаются медленно. Борозды и извилины начинают формироваться только с 11 — 12-й недели. Первыми появляются латеральная и гиппокампова извилины. Затем формирование борозд протекает очень быстро. К моменту рождения существуют все основные извилины.
Формирование коры в ходе внутриутробного развития ребенкаУченые из Института Салка (США) смогли выяснить, как в ходе внутриутробного развития формируется кора головного мозга. Ранее нейробиологи знали, что для образования коры, состоящей из шести разных слоев, требуется согласованное перемещение стволовых клеток, а также строгое соблюдение порядка их деления с образованием нескольких разных типов нейронов.. Однако, как этот порядок поддерживается, было неясным, а значит, невозможно было говорить о целенаправленном восстановлении поврежденного мозга, успешной терапии болезни Альцгеймера стволовыми клетками или о перспективах генной терапии.
В серии экспериментов ученым удалось выявить очередность перемещений и делений клеток, которую обеспечивает выявленный исследователями ген Lhx2. В течение короткого времени этот ген вместе с еще несколькими другими играет роль регулятора, задавая дальнейшее развитие клеткам и определяя, как именно будет выстроена нервная ткань.
Ученые исследовали неокортекс – область коры головного мозга, насчитывающую шесть разных слоев, в каждом из которых помимо нервных волокон расположены нейроны определенного типа. Кстати, кора мозга, которая отвечает за обоняние, также отличается непростым строением, хотя и состоит всего из трех слоев – правда, возникли они в ходе эволюции намного раньше.
Оказалось, что в неокортексе, кроме шести слоев можно выделить еще и вертикальные колонки, объединяющие несколько тысяч клеток в отдельную сеть – эти колонки соединены друг с другом определенным образом. Вся кора насчитывает миллионы нейронов даже у мышей с их крошечным, размером с некрупную вишню, мозгом, а в ходе эмбрионального развития сложнейшая сеть образуется всего из нескольких клеток.
Ученые считают колонки нейронов важнейшими образованиями коры, которые обеспечивают мышление, запоминание и некоторые другие психические функции. Чтобы лучше понять работу колонок, группа нейробиологов сейчас строит самую сложную и масштабную в истории компьютерную модель искусственного мозга.
Генные чертежи
Именно сложностью структуры коры и упорядоченностью в ней межклеточных связей обусловлены генетические «инструкции» по ее сборке. Да и гены, как подчеркивают нейробиологи, не содержат точных инструкций процесса образования коры, а обеспечивают скорее общие направления развития. Ген Lhx2 не является исключением. В определенный момент, когда стволовые клетки занимают свое положение в будущей коре, он включается и задает дальнейшее направление их развития – превращение в те или иные типы нейронов. Просто включения гена Lhx2, однако, недостаточно. Ведь его активность сообщает клеткам лишь то, что настало время приобретать ту или иную специализацию. Но вот какую? Отчасти зависит это от собственной активности гена Lhx2, но также важная роль отводится еще одному механизму, задействованному в процессе образования коры мозга. Как выяснилось в ходе исследования, сначала в клетках, из которых в будущем получится кора, активируется другой ген, Emx1. В момент, когда клетки начинают свое деление, выстраиваясь в еще несколько одинаковых слоев, начинает свою работу Lhx2. Кодируемый им белок при этом синтезируется в разных количествах, а ранее однородные слои с этого момента начинают приобретать специфические черты.
Более того, количество выработанного клеткой белка как раз и определяет дальнейшую судьбу, в каждом из слоев запускается своя цепочка биохимических реакций, зависящая от изначальной активности гена Lhx2.
Для расслоения стопки одинаковых клеток используется такой простой показатель, как количество кодируемого всего одним геном белка, в зависимости от чего клетка направляется по тому или иному пути развития.
Связь мозга и генов часто упоминается в сообщениях о нейробиологических открытиях. И это неудивительно, ведь, например, в головном мозге во время процесса обучения начинают работу сотни и даже тысячи разных генов. А свыше половины всех генов так или иначе связаны именно с развитием и функционированием мозга.
Как удалось продемонстрировать биологам, если ген Lhx2 начал свою работу не вовремя, то кора на положенные ей слои не разделяется. Промедление всего лишь на сутки приводит к тому, что вместо неокортекса у мышей развивается обонятельная, трехслойная, и более примитивная кора, а у людей подобные аномалии вообще могут привести к тяжелейшим врожденным дефектам развития мозга.
Впрочем, ученых интересовало не отсутствие коры мозга, а, как, как сообщается в пресс-релизе, выпущенном Институтом Салка, возможность замены стволовыми клетками поврежденных инсультом или нейродегенеративными болезнями участков мозга.
Первичные, вторичные и третичные поля корыОсобенности строения и функционального значения отдельных участков коры позволяют выделить отдельные корковые поля. Различают три основные группы полей в коре: первичные, вторичные и третичные поля.
Первичные поля связаны с органами чувств и органами движения на периферии, они раньше других созревают в онтогенезе, имеют наиболее крупные клетки. Это так называемые ядерные зоны анализаторов, по И. П. Павлову. Эти поля осуществляют анализ отдельных раздражений, поступающих в кору от соответствующих рецепторов. При разрушении первичных полей возникают так называемая корковая слепота, корковая глухота и т. п. Рядом расположены вторичные поля, или периферические зоны анализаторов, которые связаны с отдельными органами только через первичные поля. Они служат для обобщения и дальнейшей обработки поступающей информации. Отдельные ощущения синтезируются в них в комплексы, обусловливающие процессы восприятия. При поражении вторичных полей сохраняется способность видеть предметы, слышать звуки, но человек их не узнает, не помнит их значения. Первичные и вторичные поля имеются и у человека, и у животных. Наиболее далеки от непосредственных связей с периферией третичные поля, или зоны перекрытия анализаторов. Эти поля есть только у человека. Они занимают почти половину территории коры и имеют обширные связи с другими отделами коры и с неспецифическими системами мозга. Основным клеточным элементом здесь являются звездчатые нейроны. Третичные поля находятся в задней половине коры — на границах теменных, височных и затылочных ее областей и в передней половине — в передних частях лобных областей. В этих зонах оканчивается наибольшее число нервных волокон, соединяющих левое и правое полушария, поэтому роль их особенно велика в организации согласованной работы обоих полушарий. Третичные поля созревают у человека позже других корковых полей, они осуществляют наиболее сложные функции коры. Здесь происходят процессы высшего анализа и синтеза. В третичных полях на основе синтеза всех афферентных раздражений и с учетом следов прежних раздражении вырабатываются цели и задачи поведения. Согласно им происходит программирование двигательной деятельности. Развитие третичных полей у человека связывают с функцией речи. Мышление (внутренняя речь) возможно только при совместной деятельности анализаторов, объединение информации от которых происходит в третичных полях.
Заключение
Таким образом, каждый двигательный акт, каждое так называемое произвольное, волевое движение детерминировано, обусловлено раздражениями, поступающими в кору головного мозга из внешней или внутренней среды. Процесс развития коры головного мозга в филогенезе и онтогенезе идет сходным принципам.
Библиография
1. Воронова Н.В. Анатомия центральной нервной системы: Учебное пособие для студентов вузов — М.: Аспект Пресс, 2005. — 128 с.
2. Горелов А.А. Концепции современного естествознания : Учеб. Пособие для вузов. – М.: ВЛАДОС, 1999. – 512 с.
3. Жуков Н.И. Проблема сознания. – Минск: Университетское, 1987. – 207 с.
4. Козлов В.И. Анатомия нервной системы — М.: МИР, 2006. — 208 с.
5. Куприянович Л.И. Резервы улучшения памяти: Кибернетические аспекты.-М.: Наука, 1970.-141с.
6. Орбели Л.А., Вопросы высшей нервной деятельности, М.— Л., 1949;
7. Павлов И.П., Двадцатилетний опыт объективного изучения высшей нервной деятельности животных, Полн. собр. соч., 2 изд., т. 3, кн. 1—2, М., 1951;
Размещено на http://www.