Гены идентичности цветковых меристем
Федеральное государственное
образовательное учреждение
высшего профессионального образования
«СИБИРСКИЙ ФЕДЕРАЛЬНЫЙ УНИВЕРСИТЕТ»
Институт фундаментальной биологии и биотехнологии
ГЕНЫ ИДЕНТИЧНОСТИ ЦВЕТКОВЫХ МЕРИСТЕМ
(реферат)
Студент:
Н. Толстоноженко,
гр. ББ09-04М
Преподаватель:
к.б.н., доцент
Н.П. Белоног
Красноярск 2009
Оглавление
Глава 1. Разнообразие генов, регулирующих процесс цветения растений
1.1 Гены LFY (арабидопсиса) и FLO (львиного зева)
1.2 Гены API (арабидопсиса) и SQUA (львиного зева)
1.3 Экспрессия генов идентичности цветковых меристем
Глава 2. Молекулярная характеристика генов, контролирующих идентичность цветковой меристемы
2.1 Молекулярная характеристика генов FLO (львиный зев) и LFI (арабидопсис)
2.2 Молекулярная характеристика генов SQUA (львиный зев) и AP1 (арабидопсис)
2.3 Молекулярная характеристика гена CAL (арабидопсис)
2.4 Молекулярная характеристика гена FIM (львиный зев)
2.5 Молекулярная характеристика генов CEN (львиный зев) и TFL1 (арабидопсис)
Вывод
Глава 1. Разнообразие генов, регулирующих процесс цветения растений
Основное событие при индукции цветка - это активация небольшой группы генов, которые необходимы для превращения генеративной меристемы в цветковую меристему. У мутантов по этим генам вместо цветков развиваются побеги, побегоподобные структуры или цветки с чертами побегов. Для арабидопсиса к таким генам идентичности цветковых меристем (англ. flower meristem identity genes) относятся гены: LEAFY (LFY), APETALA1 (API), CAULIFLOWER (CAL), APETALA2 (AP2), UNUSUAL FLORAL ORGANS (UFO) и некоторые другие (табл. П.З). Для львиного зева это гены FLORICAULA (FLO), SQUAMOSA (SQUA), FIMBRIATA (FIM).
1.1 Гены LFY (арабидопсиса) и FLO (львиного зева)
Мутации по гену арабидопсиса LEAFY (LFY) обычно обладают наибольшим фенотипическим проявлением. Даже самые слабые аллели lfy превращают базальные цветки во вторичные генеративные побеги, которые образуются в пазухах небольших околоплодных листьев, похожих на прицветники. Образующиеся позднее цветки характеризуются несколькими признаками побегов; их органы наружной мутовки напоминают околоплодные листья, в пазухах которых иногда образуются вторичные цветки, расстояние между мутовками в них увеличено, расположение органов цветка близко к спиральному. У мутантов изредка образующиеся аномальные цветки состоят из наружной мутовки чашелистиков и внутренних мутовок чашелистикоподобных и карпелоподобных органов. Лепестки и тычинки у таких цветков отсутствуют, т.е. LFY играет роль не только в определении судьбы цветковой меристемы, но и в возникновении лепестков и тычинок.
Гомологом гена LFY у львиного зева является ген FLORICAU-LA (FLO). У мутантов flo на месте цветков формируются генеративные побеги. Они отличаются от цветков четырьмя основными признаками:
1)органы цветка заменяются прицветниками;
2)увеличивается длина междоузлий;
3)расположение органов близко к спиральному;
4)меристема характеризуется недетерминированным ростом.
Так проявляется одно из различий между этими дальнеродственными видами двудольных растений: фенотипический эффект мутации flo у львиного зева выражен гораздо сильнее, чем мутации lfy у арабидопсиса. Таким образом, у арабидопсиса должен существовать ген, способный частично дублировать функцию гена LFY.
Если мутантные по FLO растения долго культивировать в маленьком горшке, иногда у них развиваются терминальные структуры с карпелоподобными органами. Значит, у мутантов flo иногда могут быть восстановлены некоторые аспекты программы развития цветка, но только некоторые, так как, например, формирование лепестков и тычинок у таких мутантов обнаружить не удавалось. Цветки львиного зева, как и большинства двудольных растений, образуются в пазухах видоизмененных листьев - прицветников. У цветков арабидопсиса дикого типа прицветников нет, а у аномальных цветков мутантов lfy - есть. Мутанты lfy и flo, как и многие другие, о которых пойдет речь в этой главе, являются:
1) гомеозисные мутанты, поскольку их фенотип выражается в замене одной структуры цветка на другую, - генеративный, или соцветный, побег;
2) гетерохронические мутанты, поскольку они приводят к повторению более ранней программы развития растения, иначе говоря, к временным изменениям в процессе развития.
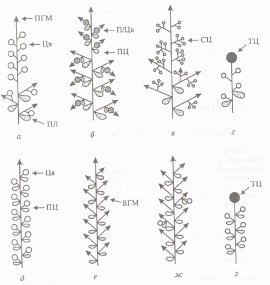
Рис. 1. Схема мутантных соцветий и соцветий дикого типа арабидопсиса (а-г) и львиного зева (д-з): а - соцветие арабидопсиса дикого типа; б - соцветие мутанта Ify; в - соцветие мутанта apl; г - соцветие мутанта tfl1; д - соцветие львиного зева дикого типа; е - соцветие мутанта flo; ж - соцветие мутанта sgua; з - соцветие мутанта cen; ПГМ - по беговая генеративная меристема; Цв - цветок; ПЦ - прицветник; П/Цв - прицветник/цветок; СЦ -соцветие; ГЦ - терминальное соцветие; ВГМ - внутренняя генеративная меристема; ЛЛ – прилистник
1.2 Гены API (арабидопсиса) и SQUA (львиного зева)
У мутантов арабидопсиса по гену APETALA1 (API) по крайней мере несколько базальных цветков замещается генеративными побегами. В основании таких генеративных побегов не формируется лист, как это характерно для побегов дикого типа. Апикальные цветки мутантов ap1 заменяются сложными детерминированными структурами, в которых на одном цветоносе развивается несколько цветков. Происходит это следующим образом: в осях органов первой мутовки (чашелистиков, вместо которых развиваются листообразные органы) возникают вторичные цветковые меристемы, которые формируют вторичные цветки. В осях органов первой мутовки таких вторичных цветков в свою очередь образуются новые цветочные меристемы - третичные или более высокого порядка, тоже дающие начало новым цветкам. Лепестки у мутантов apl обычно не образуются.
Итак, оба гена API и LFY необходимы для превращения генеративной меристемы в цветковую и обладают вырожденными, т. е. частично взаимозаменяемыми, активностями. Подтверждается это и фенотипом двойных мутантов apl lfy, у которых все цветочные примордии развиваются как генеративные побеги. Более того, фенотипические различия между слабыми и сильными аллелями как Ify, так и apl, исчезают у двойных мутантов.
Гомологом гена API у львиного зева является ген SQUAMOSA (SQUA). Как и в случае пары генов LFY/FLO, мутации squa обычно обладают более выраженным фенотипом, чем apl. На месте цветков у них, как правило, развиваются генеративные побеги, хотя изредка формируются и цветки, варьирующие по фенотипу от почти нормальных до крайне измененных. Как и для двойных мутантов apl Ify, аллели flo значительно усиливают фенотип мутантов squa, и двойные мутанты иногда полностью теряют способность формировать цветки.
Из подобных наблюдений можно заключить, что гены идентичности меристем у арабидопсиса и львиного зева функционируют сходным образом, хотя в их экспрессии и функции имеются некоторые важные различия. Активность генов идентичности меристем и их взаимодействие с продуктами других генов могут варьировать с возрастом, при изменении условий окружающей среды, а также различаться для разных видов растений. В результате этого даже одна и та же мутация может приводить к развитию растений с различающимися фенотипами. Например, карпелоподобные структуры у мутантов Ify чаще формируются на поздних этапах развития соцветия и у мутантов, культивируемых при длинном дне. Карпелоидные структуры у мутантов По также изредка образуются только у очень старых растений.
1.3 Экспрессия генов идентичности цветковых меристем
Хотя экспрессия генов FLO/LFY и SQUA/AP1 может быть индуцирована независимо друг от друга, очевидно, что наличие активности только одного из этих генов недостаточно для развития нормальных цветков. Однако оставалось неясным, достаточно ли индукции активности одного из этих генов для того, чтобы вызвать нормальное развитие цветка у растений дикого типа. Ответ на этот вопрос был получен путем конструирования трансгенных растений, в которых химерные гены, состоящие из кодирующих последовательностей генов LFY или API и конститутивного промотора p35S вируса мозаики цветной капусты (ВМЦК), экспрессируются во всех тканях, включая меристемы побегов. У таких трансгенных растений в местах, где у растений дикого типа развиваются побеги и эктопические цветки, которые по фенотипу очень близки к цветкам дикого типа. Процесс развития цветка у трансгенных растений с эктопической экспрессией LFY или API зависит от присутствия другого гена пары, поскольку цветки растений с генотипом p35S::LFY apl имеют фенотип мутантов apl (вторичные цветки, меньшее количество лепестков, формирование листоподобных органов вместо чашелистиков), а цветки растений p35S::APl Ify имеют фенотип мутантов Ify (отсутствие лепестков и тычинок, спиральное расположение органов). Таким образом, каждый из этих генов способен индуцировать активность другого гена этой пары в тех местах, где в норме не наблюдается их экспрессия, для формирования нормальных цветков необходимо комбинирование активностей обоих этих генов.
На фенотип мутантов apl влияет ген арабидопсиса CAULIFLOWER (CAL). У двойных мутантов cal «цветки» похожи на структуры, развивающиеся вместо единичных цветков у цветной капусты Brassica oleracea var botrytis (отсюда происходит и название гена - CAULIFLOWER). Клетки, которые обычно дают начало цветковой меристеме, у таких мутантов ведут себя как клетки генеративной меристемы и формируют множество дополнительных меристем, расположенных спирально. Гомозиготы по мутациям cal фенотипически не отличаются от растений дикого типа, что свидетельствует о способности гена API заменять утраченную активность CAL. Цветковые меристемы, которые формируются у мутантов apl, ведут себя как генеративные меристемы у двойных мутантов apl cal, что свидетельствует о способности продукта CAL заменять API при формировании цветковых меристем. И наоборот, поскольку органы первой и второй мутовок у мутантов apl не способны развиваться нормально, продукт CAL не может заменять API при формировании чашелистиков и лепестков. Таким образом, функции продуктов API и CAL частично вырождены, или взаимозаменяемы. В то же время мутации cal не влияют на фенотип мутантов Ify или двойных мутантов apl Ify. Пока еще не ясно, есть ли гомолог гена CAL у львиного зева.
Еще несколько генетических локусов по крайней мере немного влияют на судьбу цветковой меристемы. Хотя мутанты по гену арабидопсиса APETALA2 (АР2) и имеют цветки без признаков генеративного соцветия, в двойных мутантах apl ap2, а также Ify ар2 проявляется роль гена АР2 в формировании цветковой меристемы. Более того, при совмещении apl и ар2 в двойных мутантах наблюдается синергидный эффект; органы в мутантных цветках располагаются по спирали, что характерно для побегов, а количество цветков, развивающихся на одном цветоносе, еще увеличивается. Вероятно, АР2 может функционировать в процессе развития меристем при отсутствии активностей LFY или API.
Ген арабидопсиса UNUSUAL FLORAL ORGANS (UFO), как и LFY - важный активатор генов идентичности органов цветка. Мутанты ufo имеют черты как мутантов по генам идентичности цветковых меристем, так и генов идентичности органов цветка. Первые несколько цветков у них заменяются вторичными соцветиями. Как вторичные соцветия, так и основное соцветие у них заканчиваются карпелоидными или сепаяоидными (чашелистикообразными) структурами. Цветки таких вторичных соцветий характеризуются различными типами гомеозисных изменений в первых трех мутовках органов. Наиболее сильно при этом изменяются органы второй и третьей мутовок, в которых лепестки и тычинки заменяются химерными органами, состоящими из тканей чашелистиков, тычинок, карпел, а также филаментозных элементов. В основании многих цветков формируются прицветники или филаментозные структуры. Кроме того, мутанты ufo характеризуются уменьшением количества органов второй и третьей мутовок. Мутовчатое расположение органов у них выражено менее четко, и иногда трудно сказать, к какой мутовке относится тот или иной орган. Кроме того, у мутантов ufo нередко формируются прицветники без образования побегов или цветков в их пазухах. Разнообразие фенотипа мутантов свидетельствует о множественности функций гена UFO в развитии генеративной и цветковой меристем. Гену UFO приписываются следующие функции:
· установление кругового, или мутовчатого, характера формирования органов цветка;
· участие в контроле детерминированности развития цветка;
· активация генов типа органов цветка APETALA3 и PISTILA-ТА (см. далее), необходимых для развития лепестков и тычинок;
· определение границ между мутовками органов внутри цветочного зачатка.
Мутации в гене LFY в основном эпистатируют над мутациями ufo, что свидетельствует о возможном участии обоих генов в одном пути передачи сигнала. Чтобы проверить, справедливо ли предположение о том, что ген UFO может являться медиатором между геном идентичности меристем LFY и геном типа органов цветка АРЗ, растения арабидопсиса были трансформированы конструкциями с химерным геном из кодирующей последовательности гена UFO, слитой с конститутивным промотором p35S ВМЦК. В цветках таких трансгенных растений инициация экспрессии гена АРЗ происходила преждевременно и в местах, где у дикого типа этот ген не экспрессируется.
Таким образом, ген UFO и в самом деле является регулятором гена АРЗ. В цветках растений p35S::UFO Ify не формируются нормальные лепестки и тычинки. Ген UFO, по-видимому, способен нормально функционировать только при наличии активности гена LFY. Более вероятно, что ген UFO является частично вырожденным корегулятором, действующим вместе с геном LFY, а не просто медиатором между генами идентичности меристем и генами типа органов цветка.
Мутанты ufo похожи на мутанты по гену львиного зева FIM-BRIATA (FIM). Вероятно, эти гены являются гомологами. Фенотип мутантных по гену FIM цветков зависит от силы мутантной аллели. Цветки fim имеют лепестки с полосами тканей чашелистиков больших или меньших размеров. Такие полосы чаще всего располагаются в срединной части нижних лепестков. Иногда в пазухах второй мутовки чашелистиков развиваются вторичные цветки. В цветках сильного мутанта fim 62 тычинки часто заменяются карпелами, сливающимися с карпелами четвертой мутовки. Крайнее выражение мутантного фенотипа fim 62 - недетерминированные цветки, состоящие из чашелистиков. Мутанты по генам UFO и FIM формируют цветки, очень сильно варьирующие по фенотипу, что справедливо для всех аллелей обоих генов.
Мутации в локусе арабидопсиса TERMINAL FLOWER1 (TFL I) приводят к фенотипу, противоположному таковому для мутаций по генам идентичности цветковой меристемы. Апикальная и латеральная меристемы соцветия у них заменяются цветковыми меристемами. Кроме того, как тоже упоминалось ранее, мутанты tfll цветут раньше растений дикого типа. В отличие от растений арабидопсиса дикого типа, имеющих недетерминированное соцветие с множеством цветков и вторичными и третичными соцветиями, мутанты tfll имеют детерминированное первичное соцветие всего лишь с несколькими цветками и не образуют вторичных соцветий. Соцветие заканчивается аномальным цветком, состоящим из двух-трех неполных цветков. Похоже, вторичные меристемы соцветия у мутантов tfll заменяются меристемами цветка, а первичная генеративная меристема превращается в 2—3 смежные цветковые меристемы. Можно предположить, что ген TFL1 необходим для поддержания функциональности, или компетентности, цветковой меристемы, хотя, по-видимому, он активен и в вегетативной фазе развития. TFL1 также играет роль в негативной регуляции экспрессии генов LFY и API в апикальных и латеральных меристемах, хотя такая регуляция может осуществляться и опосредованно.
Гомологом гена TFL1 у львиного зева является ген CENTRO-RADIALIS (CEN). В норме соцветия львиного зева, как и арабидопсиса, являются недетерминированными: цветки образуются в латеральных положениях, и никогда не формируется терминальный, или апикальный, цветок, который был бы последним цветком в соцветии. При этом гены FLO, LFY, SQUA и API экспрессируются в латеральных меристемах, но не в апикальной части соцветия. Однако в апексах меристемы мутантов сеп, как и tfll, образуются терминальные цветки. Как и ожидалось, вышеперечисленные гены экспрессируются в апексах меристем этих мутантов. Более того, условия окружающей среды, которые усиливают фенотип Ify, например короткий день, ослабляют фенотип tfll. Таким образом, гены CEN и TFL1, по-видимому, являются антагонистами генов, индуцирующих развитие цветка, и, возможно, могут предотвращать их экспрессию в апексе соцветия. На основе анализа фенотипов двойных мутантов предполагают, что TFL1 является антагонистом генов LFY, API и АР2. Мутации Ify частично супрессируют tfll, двойные мутанты Ify apl или Ify ap2 характеризуются более выраженным супрессирующим эффектом на фенотип tfll. Терминальный цветок мутантов сеп обладает радиальной симметрией: все его лепестки напоминают вентральные лепестки цветков дикого типа. Количество органов цветка сеп и их филотаксис (расположение} очень вариабельны
Глава 2. Молекулярная характеристика генов, контролирующих идентичность цветковой меристемы
2.1 Молекулярная характеристика генов FLO (львиный зев) и LFI (арабидопсис)
Ген львиного зева FLO был клонирован одним из первых генов, влияющих на развитие цветка. Для его клонирования использовали мутант По-613, образующий генеративные соцветия вместо цветков. Однако гомозиготы по flo-613 изредка формировали нормальные цветки. Из семян этих цветков развивались растения дикого типа. Значит, мутация flo-613 генетически нестабильна и иногда ревертировала к аллели дикого типа. Реверсия коррелировала с эксцизией транспозона ТатЗ и, следовательно, мутантный фенотип был обусловлен инсерцией этого мобильного элемента в ген FLO. Таким образом, ген FLO был клонирован с использованием последовательности транспозона ТатЗ в качестве пробы. У всех ревертантов к дикому типу при использовании клонированного фрагмента гена FLO как пробы для гибридизации появлялся фрагмент длиной 4т. п. о. Такой же длиной обладал гибридизовавшийся с FLO фрагмент исходного растения дикого типа, использованного для транспозонового мутагенеза. При этом у некоторых ревертантов сохранялся фрагмент длиной 7.5 т. п. о., такой же, как и у мутанта flo-613. Вероятно, среди ревертантов были как гетерозиготные, так и гомозиготные растения.
Однако при гибридизации ДНК мутантных растений среди них тоже были выявлены гетерозиготы, у которых присутствовали оба фрагмента. Так, при секвенировании сайта эксцизии ТатЗ из гетерозиготы с фенотипом По обнаружили инсерцию 8 пар оснований. Таким образом, неточная эксцизия ТатЗ приводила к изменению рамки считывания, и мутантный фенотип сохранялся.
В сайте эксцизии у ревертанта отмечены замены двух пар оснований, и не обнаружены инсерции или делеции нуклеотидов. В потомстве от самоопыления всех гетерозиготных ревертантов имеются растения дикого типа и мутанты в соотношении 3:3. Потомство большинства гомозиготных ревертантов состояло только из растений дикого типа.
Мутация flo-613, как и многие другие гомеозисные мутации львиного зева, использованные для клонирования генов развития цветка и о которых пойдет речь в этой главе, была получена в массивном эксперименте по транспозоновому мутагенезу. Растения львиного зева с высокоактивными мобильными элементами Тат (от Transposone of Antirrhinum majus) выращивали при температуре 15 °С. Именно при этой температуре для мобильных элементов группы Тат наблюдается наибольшая частота транспозиций. Многие из 26000 потомков Ml этих растений, полученных при самоопылении, содержали рецессивные мутации в гетерозиготном состоянии. Такие рецессивные мутации можно было выявить в следующем поколении М2 после самоопыления растений Ml. И в самом деле, среди 80000 растений М2 было обнаружено более 15 независимых гомеозисных мутаций, затрагивающих развитие цветка (рис.2).
Ген арабидопсиса LFY был клонирован по гомологии с геном львиного зева FLO. В отличие от многих других гомеозисных генов, затрагивающих развитие цветка, FLO и LFY, похоже, уникальные гены и не являются членами генного семейства. Кодируемые ими белки были на 70 % гомологичны друг другу и не обладали значительной гомологией ни с одним из известных на момент их клонирования белков. Наличие богатого пролином домена на N-конце и кислой области в центральной части белка свидетельствуют о том, что, вероятно, они могут служить активаторами транскрипции. Подтверждается это и тем, что белок Lfy локализован в основном в ядре. Характер их экспрессии очень похож. РНК FLO и LFY начинает накапливаться в закладывающихся зачатках цветка на границе цветочной меристемы и не обнаруживается в генеративной меристеме. В момент приобретения цветочными меристемами морфологических различий (стадии 1 и 2) экспрессия усиливается в равной степени по всему молодому зачатку цветка. На стадиях 3 и 4, когда индуцируется экспрессия генов идентичности органов, РНК FLO и LFY уже практически не обнаруживается в центре начинающего формироваться цветка. Из известных генов они первыми начинают экспрессироваться в клетках, из которых затем сформируется цветок.
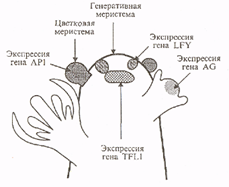
Рис.2. Схематическое изображение генеративного побега арабидопсиса и начальные этапы экспрессии генов, регулирующие развитие цветка.
Однако в характере экспрессии FLO и LFY есть одно важное отличие, помогающее понять, как развиваются меристемы. Различие является следствием того, что у цветка львиного зева есть прицветники, а у цветка арабидопсиса их нет. Ген FLO экспрессиру-ется как в примордиях прицветников, так и в цветковых меристемах. В то же время для гена LFY не наблюдается экспрессия в клетках, окружающих цветковые меристемы, и, вероятно, соответствующих клеткам львиного зева, из которых образуются прицветники. У мутантов Ify на месте цветков дикого типа без прицветников образуются генеративные побеги или «цветки» с околоплодными листьями, которые можно отождествлять с прицветниками. Значит, продукт LFY дикого типа подавляет инициацию или образование прицветников. Предполагают, что ген LFY влияет на судьбу группы клеток на периферии цветочной меристемы: в цветках дикого типа все или большинство этих клеток формируют цветочную меристему, тогда как у мутантов Ify эти клетки дают начало меристеме и примордию прицветника. В этом отношении мутант Ify похож на дикий тип растений таких видов, как Antirrhinum majus, у которых из одного зачатка образуются меристема цветка и прицветник. Считается, что наличие прицветников - эволюционно более древний признак.
Насколько универсальны охарактеризованные механизмы индукции цветка? Пока что на этот вопрос ответить не просто. Похоже, что для табака, который является более близким родственником львиного зева, чем арабидопсис, индукция развития цветковой меристемы происходит по-другому. Гены табака Nicotiana tabacum NFL1 и NFL2, гомологи генов FLO и LFY, транскрибируются во время как вегетативного, так и репродуктивного развития растений. Их экспрессия не инициируется во время индукции цветка, как это характерно для генов FLO и LFY. Возможно, это связано с тем, что табак имеет терминальный цветок, в то время как соцветия львиного зева и арабидопсиса не детерминированы. Трансгенные растения арабидопсиса, в которых конститутивно экспрессируется ген LFY, тоже имеют терминальный цветок. В геномах диплоидных растений львиного зева и арабидопсиса имеется только одна копия гена FLO или LFY. Табак же является аллотет-раплоидом, и каждый из его диплоидных геномов содержит по одной копии гомологичного FLO/LFY гена.
2.2 Молекулярная характеристика генов SQUA (львиный зев) и AP1 (арабидопсис)
Ген львиного зева SQUA был клонирован с использованием в качестве пробы гена DEF из этого же растения, содержащего MADS-бокс (MADS-бокс — это последовательность ДНК, кодирующая домен MADS. Он получил свое название от четырех клонированных первыми генов с этим боксом: МСМ1, AGAMOUS, DEFICIENS, SRF. Домен MADS представляет собой консервативную область белков - факторов транскрипции, участвующую в связывании с ДНК (подробнее об этом см. далее). По гомологии с DEF было выделено 9 независимых генов львиного зева. Один из клонов при гибридизации геномных блотов по Саузерну проявлял рестрикционный полиморфизм для ДНК дикого типа и нестабильной мутации squa-29. Ревертанты SQUA имели фрагмент ДНК такой же длины, как и растения дикого типа.
Ген арабидопсиса API был клонирован как новый член семейства генов, содержащих MADS-бокс.
Белки Squa и Apl характеризуются 68%-ной гомологией. Наличие в этих белках домена MADS предполагает, что они могут специфически связываться с ДНК и функционировать как активаторы транскрипции. РНК SQUA и API индуцируется в молодых цветочных примордиях, как только они становятся видимыми на границе генеративной меристемы (чуть позже, чем РНК FLO и LFY). На стадиях I и 2 РНК SQUA и API однородно экспрессируется по всему примордию цветка (рис. 11.33), на стадии 3 экспрессия снижается в двух внутренних мутовках, и затем РНК API обнаруживается только в примордиях чашелистиков и лепестков. РНК SQUA, кроме того, менее интенсивно экспрессируется и в примордиях прицветников и карпел. В вегетативных органах, за исключением самых верхних листьев, примыкающих к соцветию, РНК SQUA не обнаруживается (рис. 11.32, в).
Ген SQUA экспрессируется в мутантах flo, а ген FLO — в мутантах squa. Таким образом, FLO и SQUA активируются независимо друг от друга. Подобным образом, ген LFY экспрессируется в мутантах apl. Фенотип двойных мутантов flo squa и Ify apl выражен значительно сильнее, чем фенотип одиночных мутантов по этим генам, что также свидетельствует о независимом и синергид-ном действии этих генов в процессе формирования цветковой меристемы и о частичной вырожденности их активностей.
Ген LFY не только необходим для формирования цветка, но и достаточен для определения идентичности цветка, т.е. развития меристемы как цветковой меристемы. Основной побег трансгенных растений p35S::LFY с конститутивной экспрессией этого гена переходит к цветению раньше, чем растения дикого типа, а вторичные побеги превращаются в цветки. Хотя ген API экспрессируется несколько позднее, чем LFY, он тоже необходим для инициации развития цветка. Конститутивная экспрессия трансгена p35S::APl тоже ускоряет переход растения к цветению и приводит к образованию цветков на месте вторичных побегов. По-видимому, гены LFY/AP1 и FLO/SQUA оказывают наибольшее влияние на инициацию цветковых меристем. Однако в этот процесс вносят определенный вклад и другие гены.
2.3 Молекулярная характеристика гена CAL (арабидопсис)
Ген арабидопсиса CAULIFLOWER очень похож на ген API, что и было использовано для его клонирования. Как и API, ген CAL тоже кодирует белок с доменом MADS, и, следовательно, тоже может являться активатором транскрипции. Последовательности белков Cal и Apl имеют гомологию 70 %. Внутри домена MADS они различаются только пятью аминокислотными остатками из 56, т. е. оба белка, вероятно, узнают похожие последовательности ДНК и участвуют в регуляции сходных генов. Таким образом, функциональная вырожденность этих генов, обнаруживаемая в генетических исследованиях, подтверждается их структурным сходством. Однако, хотя данные молекулярного анализа хорошо согласуются с фактом функциональной вырожденности генов CAL и API, они не дают ответа на вопрос, почему CAL может только частично заменять API.
Ген CAL начинает экспрессироваться в примордиях цветков одновременно с геном API, и затем тоже обнаруживается только в примордиях двух наружных мутовок органов. В отличие от API, РНК которого накапливается в больших количествах в процессе развития лепестков и чашелистиков, РНК CAL в примордиях этих органов обнаруживается лишь в небольших количествах. Возможно, именно поэтому CAL неспособен заменить API при формировании лепестков и чашелистиков. Какова же роль гена CAL в развитии цветка? Данные молекулярного анализа свидетельствуют о том, что в двойных мутантах apl cal концентрация РНК LFY очень сильно редуцирована, в то время как в мутантах apl она не отличается от таковой для дикого типа. Таким образом, функция CAL может заключаться в поддержании активности гена LFY. Вероятно, поэтому и фенотип тройных мутантов Ну apl cal не отличается от фенотипа двойных мутантов Ify apl: если ген LFY инактивирован, ген CAL уже неспособен влиять на его активность.
Соцветия цветной капусты Brassica oleracea var. botrytis и капусты-брокколи Brassica oleracea var. italica представляют собой одну из аномалий развития цветка. Как и многие другие разновидности используемой для еды капусты - такие, как капуста белокочанная и краснокочанная, брюссельская, листовая, - обе эти разновидности тоже произошли от Brassica oleracea дикого типа. Как известно, Brassica и арабидопсис относятся к одному семейству - крестоцветных, и их соцветия дикого типа и фенотипа cauliflower морфологически очень похожи. При исследовании двух достаточно отдаленных видов растений, арабидопсиса и львиного зева оказалось, что механизмы, контролирующие развитие цветка, у них довольно консервативны. Гомологи почти всех клонированных генов есть у обоих видов, и типы их экспрессии очень похожи. Следовательно, не должно быть сюрпризом то, что молекулярная основа фенотипа cauliflower у арабидопсиса и близкородственной Brassica oleracea var. botrytis могут быть идентичны. Чтобы проверить, так ли это на самом деле, были клонированы гомологи гена CAL - bobCAL из цветной капусты, boiCAL из брокколи и boCAL из Brassica oleracea дикого типа. Как оказалось, открытая рамка считывания гена bobCAL прерывается стоп-кодоном в пятом экзоне, и соответствующий мутантный белок имеет молекулярный вес 17 кДа вместо белка дикого типа с молекулярным весом 30 кДа. В гене boiCAL не обнаружили подобный стоп-кодон, однако, в нем имеется делеция кодонов пяти аминокислотных остатков, а также аминокислотные замены в двух из трех кодонах, которые изменены у мутанта арабидопсиса cal-1. Таким образом, эти данные свидетельствуют в пользу того, что фенотипы соцветий цветной капусты и брокколи по крайней мере частично обусловлены мутациями в гене CAL.
2.4 Молекулярная характеристика гена FIM (львиный зев)
Ген львиного зева FIM был клонирован с использованием метода транспозоновой мишени. Мутанты fun 619 и fim 620 с инсерцией мобильных элементов были получены в описанном выше эксперименте по мутагенезу. Ген арабидопсиса UFO был клонирован по гомологии с геном FIM: фрагмент гена FIM почти полной длины использовали как пробу для гибридизациии с геномной библиотекой арабидопсиса. Нуклеотидные последовательности генов FIM и UFO обладают значительной степенью гомологии. В то же время последовательности этих генов не обнаруживают значительного сходства с какими-либо другими нуклеотидными последовательностями, имеющимися в базах данных. Оба гена, FIM и UFO, активируются в молодых цветковых меристемах раньше генов типа органов цветка. Инициация их экспрессии наблюдается в центре цветковой меристемы, и затем эта область «расходится» в стороны и превращается в кольцо, в центре которого РНК FIM или UFO уже не обнаруживаются. Еще позднее экспрессия этих генов ограничивается небольшой областью между мутовками лепестков и других органов. UFO работает только в области между чашелистиками и лепестками, в то время как ген FIM экспресси-руется как между мутовками чашелистиков и лепестков, так и между мутовками лепестков и тычинок.
Между гомологами FIM/UFO, как и в случае другой пары FLO/LFY, обнаруживаются и другие различия. У цветков мутантов ufo, как и у мутантов Ify, образуются прицветники. Таким образом, одной из функций генов LFY и UFO является подавление образования прицветников. Подобной функцией не обладают их гомологи FLO и FIM. Ген UFO начинает действовать чуть раньше, чем ген FIM. Транскрипты UFO обнаруживаются на более ранней стадии, чем транскрипты FIM.
Мутации ufo выражаются в изменении размеров, количества и закладки органов цветка, а также в их слиянии. Это говорит о том, что ген UFO участвует в регуляции клеточных делений и пролиферации органов. Подтверждается это и фенотипом трансгенных растений арабидопсиса с геном UFO, слитым с конститутивным промотором p35S ВМЦК: в цветках таких растений развивается больше лепестков и тычинок, при этом общее количество лепестков и тычинок может быть как больше, так и меньше восьми, а количество репродуктивных органов может достигать шестнадцати (против шести в цветках дикого типа). Гены UFO/FIM характеризуются наличием F-бокса, который имеется также в генах, контролирующих синтез таких белков, как циклин А, циклин F и Cdc4, вовлеченных в контроль клеточного цикла у дрожжей. Некоторые белки с F-боксом, например MetSop и GRR1 дрожжей, функционируют как ингибиторы транскрипции. Белок Fim связывается с белками семейства Fap (от EIM-associated £roteins), которые очень напоминают белки Skpl дрожжей и человека. Последние образуют комплексы с белками, содержащими F-бокс, вызывая деградацию белков и развитие клеточного цикла. Ген FIM играет также ключевую роль в активации генов идентичности цветковой меристемы.
2.5 Молекулярная характеристика геновCEN (львиный зев) и TFL1 (арабидопсис)
Гены CEN и TFL1 активируются в зрелых соцветиях дикого типа чуть ниже верхней части апикальной меристемы соцветия (рис. 3). РНК TFL1 также обнаруживается в латеральных генеративных меристемах и по всей длине генеративного стебля. Таким образом, вероятно, работа гена TFL1 в генеративном стебле приводит к задержке перехода к цветению. Ген CEN экспрессируется в генеративном стебле очень слабо. Вероятно, поэтому мутации сеп, в отличие от мутаций tfl, не влияют на время перехода к цветению. Экспрессия гена TFL1 в апикальной меристеме ингиби-рует работу в ней генов LFY и API, а гена CEN ингибирует экспрессию гена FLO и таким образом предотвращает развитие апекса побега как цветочной меристемы (рис. 11.34). Неожиданным результатом анализа взаимодействия CEN с другими генами оказалось то, что ген FLO в свою очередь необходим для экспрессии CEN в апикальной меристеме. В мутантных по FLO растениях, у которых формируется нефункциональная РНК FLO, РНК CEN не обнаруживается.
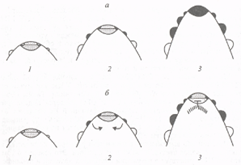
Рис. 3. Схематическое изображение начальных этапов развития генеративной меристемы мутанта сеп (а) и растений львиного зева дикого типа (б).
1 - момент закладки цветковых меристем; 2 и 3 - апекс генеративной меристемы через 2 и 6 дней с момента закладки цветковых меристем. Цветковые меристемы показаны черным цветом, вегетативные - белым, апикальные меристемы - точками. Прицветники не показаны. В мутанте cen ген FLO после некоторой задержки активируется в апикальной меристеме. В растениях дикого типа экспрессия FLO индуцирует экспрессию CEN. В субапикальной зоне (сигнал от FLO показан стрелками, зона экспрессии GEN заштрихована). Активация CEN ингибирует экспрессию FLO в апикальной меристеме.
Вывод
Гены, о которых говорилось выше, необходимы для успешного перехода к цветению. Можно выделить две группы таких генов: гены, влияющие на время индукции цветения, и гены идентичности цветковых меристем.