Выделение мембранных белков
Содержание
Введение. 2
1. Структура мембранных белков. 3
2. Выделение мембранных белков. 6
2.1 Солюбилизация мембранных белков. 7
2.1.1 Периферические белки. 7
3. Характеристика очищенных интегральных мембранных белков. 10
3.1 Определение молекулярной массы субъединиц (электрофорез в ПААГ) 10
3.2 Определение молекулярной массы нативного белка с помощью гидродинамических методов. 11
3.3 Метод радиационной инактивации. 12
3.4 Спектральные методы.. 13
3.4.1 Метод кругового дихроизма. 14
3.4.2 Инфракрасная спектроскопия и спектроскопия комбинационного рассеяния 15
3.4.3 ЯМР-спектроскопия. 16
3.5 Определение ферментативной активности. 16
3.6 Изучение стехиометрии субъединиц. 17
3.7 Изучение трехмерной структуры с помощью рентгеновской дифракции и реконструкции изображения. 18
3.7.1 Кристаллизация мембранных белков. 18
3.7.2 Реконструкция изображения и двумерные кристаллы.. 20
4. Пример структурных исследований мембранных белков. 21
4.1 Структура поринов. 21
Заключение. 24
Список литературы.. 25
Введение
Цель работы: ознакомление с методами изучения мембранных белков.
Поставленная цель раскрывается через следующие задачи:
исследовать структуру мембранных белков;
рассмотреть методы их выделения и очистки;
методы исследования характеристик мембранных белков.
Известно, что более трети структурной части генома кодирует мембранные белки, однако среди известных пространственных структур лишь менее половины процента принадлежит белкам этого класса.
При этом их роль в организме трудно переоценить. Большое количество функционально значимых белков являются мембранными: рецепторы, каналы, различные ферменты и пр. Кроме того, многие из них являются мишенями для лекарственных препаратов: более 70% существующих лекарственных средств действуют именно на мембранные белки. Таким образом, изучение механизмов функционирования мембранных белков необходимо, а одним из первых шагов к пониманию механизма является получение пространственной структуры.
1. Структура мембранных белков
Основная роль липидов в составе мембран заключается в стабилизации бислойной структуры, а белки являются активными компонентами биомембран.
Используя мембрану эритроцитов как модель, исследователи выявили два типа мембранных белков. Белки первого типа, называемые периферическими белками, связаны с мембраной в основном ионными взаимодействиями. Если обработать препарат мембран буферным раствором с высокой концентрацией солей, белки этого типа освобождаются от мембраны и переходят в буфер. Примеры периферических белков – фибронектин(локализован на наружной поверхности большинства клеток, исключая циркулирующие клетки крови) и спектрин (находится на внутренней поверхности большинства клеточных мембран, особенно в эритроцитах).
Мембранные белки второго типа называются интегральными белками. Эти протеины или погружены в толщу липидного бислоя, или пронизывают мембрану насквозь (трансмембранные белки). К интегральным относят также белки, ковалентно связанные с молекулами мембраны. Все интегральные белки можно выделить. только разрушив мембрану. Для выделения и изучения интегральных белков их очищают от липидов, либо экстрагируя их органическими растворителями (такими как ацетон или спирты), либо растворяют липиды с помощью детергентов. Большинство мембранных белков являются интегральными.
На заре развития мембранологии полагали, что мембранные белки по своей структуре довольно гомогенны и уложены в виде β-слоев по поверхности бислоя. Сейчас принято считать, что большинство мембранных белков в своей мембранной части состоят из одной или нескольких ассоциированных α-спиралей, многие мембранные белки олигомеризуются. Причем “правильная” олигомеризация α-спиралей является необходимым условием выполнения белком своей функции. Наиболее изученным из олигомеризующихся мембранных белков является белок эритроцитов человека - гликофорин А, который образует устойчивый димер не только в природных системах, но и в искусственных липидных средах, таких как мицеллы додецилфосфохолина (DPC). Интегральные мембранные белки могут оказаться гораздо сложнее, чем мы сейчас представляем. Классификация растворимых белков по типам структур была проведена только после того, как установили с высоким разрешением структуру более 100 различных белков. Что касается трансмембранных белков, то это удалось сделать только в одном случае - для белка фотосинтетического реакционного центра бактерий. Вместе с электронно-микроскопическими данными низкого разрешения о структуре бактериородопсина это единственный источник, на котором может основываться построение моделей для большинства других трансмембранных белков. Молекулярная масса мембранных белков обычно варьирует в пределах от 10 тыс. до 240 тыс.
Еще один важный момент - способы прикрепления белков к мембране:
1. Связывание с белками, погруженными в бислой. В качестве примеров можно привести F1-часть Н + - АТРазы, которая связывается с Fo-частью, погруженной в мембрану; можно упомянуть также некоторые белки цитоскелета.
2. Связывание с поверхностью бислоя. Это взаимодействие имеет в первую очередь электростатическую природу (например, основный белок миелина) или гидрофобную (например, поверхностно-активные пептиды и, возможно, фосфолипазы). На поверхности некоторых мембранных белков имеются гидрофобные домены, образующиеся благодаря особенностям вторичной или третичной структуры. Указанные поверхностные взаимодействия могут использоваться как дополнение к другим взаимодействиям, например к трансмембранному заякориванию.
3. Связывание с помощью гидрофобного "якоря"; эта структура обычно выявляется как последовательность неполярных аминокислотных остатков (например, у цитохрома 65). Некоторые мембранные белки используют в качестве якоря ковалентно связанные с ними жирные кислоты или фосфолипиды.
4. Трансмембранные белки. Одни из них пересекают мембрану только один раз (например, гликофорин), другие - несколько раз (например, лактозопермеаза; бактериородопсин).
Различиями между наружными (или периферическими) и внутренними (или интегральными) мембранными белками не задается однозначно способ их прикрепления к бислою; эти различия определяют лишь относительную силу их связывания.
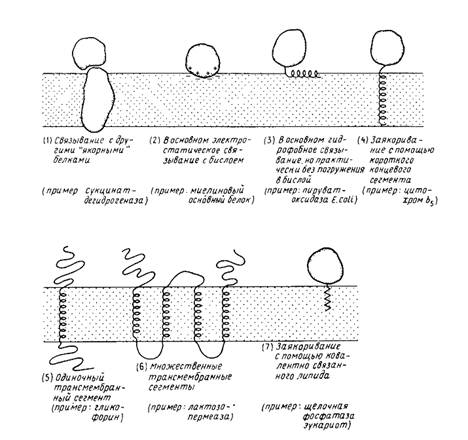
Рис. 1
Различные способы прикрепления мембранных белков к мембране. Пептидный якорь (4) может находиться либо на N-, либо на С-конце молекулы. N – и С-концы трансмембранных белков (5 и 6) могут находиться как у наружной, так и у внутренней поверхности мембраны.
2. Выделение мембранных белков
Очистка и характеристика мембранных белков ставят перед исследователем целый ряд специфических проблем, с которыми он обычно не сталкивается, работая с растворимыми белками. Мембранные белки, как правило, прочно связаны с липидным бислоем и фактически нерастворимы в воде. Поэтому для их солюбилизации и очистки приходится применять детергенты или другие разрушающие мембрану вещества. Поскольку выделение интегральных белков обычно сопряжено с разрушением мембраны, во многих случаях необходимо убедиться, что в процессе выделения и очистки белка его функциональная активность не оказалась нарушенной или утраченной. Для этого, в частности, можно попытаться провести реконструкцию,
т.е. встроить очищенный белок обратно в мембрану. Функциональную активность некоторых мембранных белков, таких, как ионные каналы или транспортные белки, можно охарактеризовать и измерить только в реконструированных мембранных системах. Для других мембранных белков, выполняющих функции ферментов или рецепторов, полезную информацию часто можно получить, используя солюбилизированные препараты. Методы солюбилизации и реконструкции не только дают ценную информацию о функциях мембранных белков; их можно также использовать для того, чтобы перевести эти белки в состояние, удобное для проведения детального структурного анализа. Несомненно, детергенты будут играть ключевую роль в совершенствовании методов кристаллизации мембранных белков для последующего рентгеноструктурного анализа.
При солюбилизации детергентами возникает вопрос о возможности избирательного извлечения из мембраны именно тех компонентов, которые интересуют исследователя. Поэтому любой новый детергент желательно проверять на возможность избирательной экстракции определенных мембранных компонентов. Ни один из имеющихся детергентов не является универсальным.
Это обусловлено тремя обстоятельствами:
1) сильными различиями в действии детергентов даже на одни и те же мембранные белки;
2) отсутствием единой стратегии солюбилизации и реконструкции;
3) сложным характером взаимодействий между молекулами белков, липидов и детергентов, имеющих столь разную химическую природу.
Существуют определенные требования к детергентам, применяемым для солюбилизации и реконструкции. Детергент должен солюбилизировать, но не денатурировать белок и должен быть легко доступен в чистом виде; желательно, чтобы детергент был недорогим.
2.1 Солюбилизация мембранных белков
2.1.1 Периферические белки
В зависимости от задачи, которая стоит перед исследователем, мембрана может быть подвергнута мягкой или жесткой обработке.
При мягких условиях обработки используют как растворы с низкой ионной силой (например, 0,1-1 мМ ЭДТА, который удаляет двухвалентные катионы), так и буферы с высокой ионной силой, содержащие NaCl и КСl в концентрации более 1 М, с добавлением ЭДТА или без нее. Не следует вводить в эти растворы такие анионы, как йодид или дииодсалицилат, поскольку они обладают хаотропнымн свойствами и могут действовать подобно детергентам. рН среды может меняться в пределах от 6,0 до 8,0. В этих условиях необратимая денатурация интегральных или периферических белков маловероятна.
При более жесткой обработке из мембран можно удалить значительные количества белка (>50% от его общего содержания в мембранах), но, с другой стороны, такая обработка обычно приводит к денатурации, во многих случаях необратимой. В качестве примера можно привести обработку мембран 6 М гуанидинийхлоридом, 8 М мочевиной, 1 мМ п-хлормер-курибензоатом, разбавленными кислотами (рН 2,0-3,0) или щелочами (рН 9,5-11,0). В кислых условиях иногда наблюдается осаждение солюбилизированных белков, и поэтому чаще прибегают к щелочной обработке.
Необходимо иметь в виду, что в результате удаления значительных количеств белка мембрана может морфологически измениться, в частности может произойти ее выворачивание или замыкание в везикулы. Поэтому следует так подобрать условия последующего центрифугирования, чтобы гарантировать полное осаждение мембран. Если используется кислотно-щелочная обработка, следует как можно быстрее вернуть рН к исходным нейтральным значениям.
2.1.2 Интегральные белки
При обработке мембран для получения интегральных мембранных белков могут высвобождаться или активироваться протеазы. Поэтому нередко на этой стадии приходится добавлять ингибиторы протеаз, даже если они уже были введены на предыдущих этапах выделения мембран. Существуют разные и довольно сложные смеси ингибиторов, которые можно использовать в зависимости от чувствительности системы к действию протеолитических ферментов. Весьма полезным, но не универсальным агентом является ингибитор сериновых протеаз, фенилметилсульфонилфторид (ФМСФ). Этот реагент хранят в концентрации 100 мМ в изопропаноле или этаноле и добавляют в инкубационную среду до концентрации 100 мкМ. Следует помнить, что он имеет довольно короткое время жизни в водных средах (ПО и 35 мин соответственно при рН 7,0 и 8,0 и 25°С). Для ингибирования SH-протеаз может оказаться полезным 10 мМ тетратионат натрия.
3. Характеристика очищенных интегральных мембранных белков
Характеристика очищенных мембранных белков, даже самых простых, может составлять определенные трудности. Как и в случае растворимых белков, нужно определить число и молекулярную массу полипептидных субъединиц, их стехиометрию, размер и, возможно, форму молекулы, а также, если это необходимо, биохимическую активность.
3.1 Определение молекулярной массы субъединиц (электрофорез в ПААГ)
Электрофорез в полиакриламидном геле в присутствии додецил-сульфата натрия - это обычная методика, но в случае интегральных мембранных белков при ее применении возникают особые проблемы. В этом методе додецилсульфат связывается с полипептидными цепями, и комплексы белок-ДНС разделяются в полиакриламидном геле в соответствии с их стоксовыми радиусами, которые в большинстве случаев зависят от молекулярной массы. Молекулярную массу определяют, сравнивая электрофоретическую подвижность данного комплекса и известного стандарта. Однако связывание ДСН с неизвестным белком может качественно отличаться от связывания со стандартами, и тогда будет получен неправильный результат. Подобная ситуация наблюдается для интегральных мембранных белков с высоким содержанием неполярных аминокислотных остатков. Возможна и другая ситуация. Связывающийся с ДСН мембранный белок может находиться в не полностью развернутом состоянии, что тоже приведет к аномальному повышению электрофоретической подвижности из-за образования более компактного комплекса белок-ДСН. Все эти эффекты весьма существенны. Например, лактозопермеаза имеет кажущуюся молекулярную массу 33 000, если измерять ее с помощью электрофореза в ПААГ в присутствии ДСН; в действительности же, как показывают результаты генетического анализа, ее молекулярная масса равна 46 000. Еще одна проблема - возможное наличие четвертичной структуры. Некоторые мембранные белки агрегируют даже в присутствии ДСН. Например, гликофорин А или белок оболочки бактериофага М13 (или fd) при электрофорезе в полиакриламидных гелях с ДСН находятся в основном в виде димеров.
Итак, оценка молекулярной массы субъединиц сильно неполярных интегральных мембранных белков, определенная с помощью электрофореза в ПААГ с ДСН, может оказаться неверной. К несчастью, простая альтернатива этому методу отсутствует, и правильную величину часто получают либо по данным о полной первичной последовательности (обычно последовательности соответствующего гена), либо с помощью точного гидродинамического анализа.
3.2 Определение молекулярной массы нативного белка с помощью гидродинамических методов
Применение этих методов для мембранных белков может быть сопряжено с большими трудностями, вызванными связыванием детергента.
Для нахождения молекулярной массы белка используют два метода.
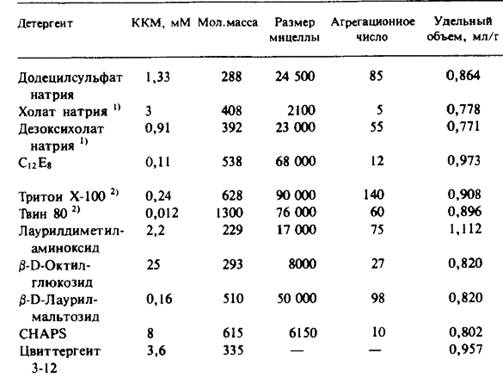
1. Прямо измеряют количество связанного детергента на 1 г белка. Для этого используют спектральные методы или радиоактивно меченный детергент, а для выделения комплексов применяют различные методы, например гель-фильтрацию. Установив относительное содержание белка и детергента в комплексе, cреднее значение парциального удельного объема получают как средневзвешенное соответствующих величин для чистого белка и чистого детергента. После этого без труда находят Mк, а поскольку соотношение между белком и детергентом в комплексе известно, находят молекулярную массу белка.
2. Измеряют S0 (коэффициент седиментации) в средах с разными значениями плотности раствора ρ. Такие среды обычно получают, используя смеси Н2О и D2O. Из графика зависимости S0 от ρ находят как Mк, так и Vк (ср). При этом предполагается, что Vк - это средневзвешенное соответствующих величин для чистого белка и чистого детергента.
Vк(ср) = (Доля белка) • V(ср) белок + (Доля детергента) • V(ср) детергент.
Оценив V(ср) белок и взяв V(ср) детергент из таблицы, получают молекулярную массу белковой составляющей Mк.
Для построения графика зависимости S0 от ρ проводят аналитическое центрифугирование. Можно проводить центрифугирование и в градиенте плотности сахарозы, используя смеси Н2О и D2O, но анализ результатов в этом случае гораздо сложнее, хотя принципиально не отличается от предыдущего случая.
3.3 Метод радиационной инактивации
Метод радиационной инактивации для определения размера мишени все чаще применяется при исследовании мембранных белков. Изучать можно как очищенные белки, так и неочищенные препараты, в том числе интактные биомембраны. Суть метода состоит в определении доли белковых молекул, получающих повреждения при облучении. Для этого используют ферментативные методы связывания гормонов или других лигандов или спектральные методы. Процедура состоит в следующем. Образец, обычно замороженный, подвергают высокоэнергетическому облучению (например, облучают пучком электронов из синхротрона). Через разные промежутки времени отбирают пробы, размораживают их и проводят измерения. Повреждения белка под действием излучения (в частности, разрыв ковалентных связей) выявляют, например, с помощью электрофореза в ПААГ с ДСН. Как показывает опыт, некоторые субъединицы полностью утрачивают биологическую активность при внесении радиационного повреждения в любое место полипептидной цепи. Ключевым моментом является то, что, чем крупнее белковая молекула, тем больше вероятность ее повреждения и, следовательно вероятность инактивации. Эта вероятность зависит не от формы молекулы, а от ее массы. Обычно для того, чтобы облегчить интерпретацию результатов, параллельно облучают белок с известной молекулярной массой. Если исследуемый белок содержит более одной субъединицы, возникают определенные трудности при анализе результатов.
3.4 Спектральные методы
Для определения содержания α - спиралей и β - слоев в мембранных белках используют несколько методов. В отсутствие трехмерной организации на их основе можно попытаться построить соответствующие модели. Чаще всего используется метод кругового дихроизма (КД). Все более широкое применение находят инфракрасная и рамановская спектроскопия, а также ЯМР.
3.4.1 Метод кругового дихроизма
Метод основан на измерении разности поглощения лево - и правополяризованного света; эта оптическая активность является мерой хиральности молекул, или мерой их асимметрии. В дальней ультрафиолетовой области (от 190 до 240 нм) КД определяется в основном поглощением амидов карбонильных групп полипептидного остова. При наличии участков вторичной структуры, например α-спиралей, спектр КД имеет вполне определенные особенности, связанные с особенностями электронного окружения амидных групп в этих структурах. Анализируя спектр КД белков, его обычно представляют как сумму компонентов, отвечающих поглощению разных участков белковой молекулы: α-спиралей, β-слоев и случайных клубков. Определив тем или иным способом спектры каждой из этих структур, производят их суммирование, подбирая соответствующие коэффициенты таким образом, чтобы было достигнуто наилучшее соответствие измеренному спектру. Подобранные весовые коэффициенты представляют собой ту долю, которая приходится в молекуле на каждый из типов вторичной структуры.
Эти методы были разработаны для растворимых белков, но нет никаких оснований сомневаться, что их можно с успехом применять и для мембранных белков. Скорее всего, у последних имеются участки с такими же типами вторичной структуры, как и у растворимых белков, и при их изучении возникнут такие же трудности. Некоторые белки можно изучать in situ, используя суспензии мембран. Примерами такого рода являются бактериородопсин из пурпурной мембраны Halobacterium halobium и Са2 + - АТР-аза из мембраны саркоплазматического ретикулума. Очищенные мембранные белки можно исследовать с помощью КД и в присутствии детергентов, если поглощение последних в дальней УФ-области не слишком велико, или в составе реконструированных везикул. Здесь возникают две проблемы:
1) дифференциальное светорассеяние, когда размер мембранных частиц гораздо больше длины волны света;
2) выравнивание поглощения из-за концентрирования белка в мембранах или везикулах, т.е. из-за негомогенности его распределения в растворе. Эти артефакты могут быть весьма существенными, однако их можно учесть с помощью соответствующих методов.
К сожалению, для внутренних мембранных белков отсутствуют структурные данные высокого разрешения, поэтому точная интерпретация спектров КД невозможна. За исключением нескольких случаев, разные спектральные методы не использовались для изучения одного и того же белка и количественное сравнение результатов не проводилось. Интересно, что для бактериородопсина, который исследовали методами КД, ИК и ЯМР, во всех трех случаях были получены одинаковые результаты, свидетельствующие о значительном содержании в этом белке β-слоев. Тем не менее у каждого метода имеются существенные недостатки. Так, данные о высоком содержании в бактериородопсине β-слоев (-46%) в значительной мере зависят от способа учета оптических артефактов. Судя по данным электронно-микроскопической реконструкции, характеризующимся относительно низким разрешением, в бактериородопсине 80% приходится на долю α-спиралей, а β-слои отсутствуют совсем. Чтобы понять причину этих несоответствий, необходимо провести структурный анализ белка с атомным разрешением.
3.4.2 Инфракрасная спектроскопия и спектроскопия комбинационного рассеяния
Эти методы не только позволяют получить сведения о конформации мембранных липидов, но и могут использоваться для исследования вторичной структуры белков. Колебательный спектр полипептидного остова зависит от типа вторичной структуры и дает информацию о содержании в молекуле α - и β - структур. Этими методами можно исследовать высушенные на воздухе пленки, водные суспензии мембран, а также очищенные белки как в присутствии детергента, так и в составе реконструированных везикул.
3.4.3 ЯМР-спектроскопия
Этот метод также может использоваться для изучения мембранных белков. Однако возможности метода в этом случае ограничены, что связано главным образом с относительно медленными движениями интегральных мембранных белков in situ и в комплексах с детергентом.
3.5 Определение ферментативной активности
Одним из наиболее важных методов характеристики очищенных мембранных белков несомненно является определение биохимической активности. При этом используются в основном такие же критерии, как и для растворимых белков, но могут возникать и свои трудности. Первая из них связана с тем, что биохимическая активность мембранных белков часто очень сильно зависит от связывания с белком липидов и детергентов. Потеря активности может быть как обратимой, так и необратимой. Целесообразно иметь какую-то оценку удельной активности исследуемого белка in vivo или в составе мембран до солюбилизации. Избыток детергента может оказывать ингибирующий эффект, например за счет разбавления неполярных субстратов в популяции мицелл и уменьшения ферментативной активности. Измеряя активность любого мембранного белка, необходимо иметь в виду, что in situ он находится в окружении липидов, обеспечивающих оптимальную активность. Вторая проблема связана с белками, обладающими "трансбислойной" активностью; примерами могут служить белки, образующие каналы, и транспортные белки. В этих случаях необходимо учитывать перемещение растворенных веществ из одного компартмента в другой (например, внутрь липосом и наружу).
3.6 Изучение стехиометрии субъединиц
Многие мембранные ферменты представляют собой комплексы, состоящие из нескольких субъединиц. В качестве примера можно привести Н + - АТРазу, Na +/ К + - АТРазу, митохондриальные комплексы электронного транспорта и фотосинтетические реакционные центры. Некоторые интегральные мембранные белки прочно связаны с растворимыми белками с помощью нековалентных взаимодействий (примерами могут служить Fo - и F1-компоненты митохондриальной Н + - АТРазы). В Е. сoli Fo-компонент, содержащий по данным электрофореза в ПААГ-ДСН три типа субъединиц (а, b, с), образует протонный канал, a F1, состоящий из пяти типов субъединиц, содержит активный центр, участвующий в гидролизе АТР. Для таких белков очень важно определить характер субъединиц, стехиометрию комплекса и ближайшие взаимодействия его компонентов. Это весьма непростая задача даже тогда, когда белковый комплекс уже изолирован. Возникающие здесь проблемы по существу не отличаются от таковых для растворимых белковых комплексов, но имеются и свои дополнительные сложности.
Прежде всего следует иметь в виду, что взаимодействие между субъединицами очень сильно зависит от типа липидов и детергентов, с которыми связаны белки.
Еще одна проблема связана с тем, что в бислое мембранные белки могут образовывать комплексы из-за высокой их локальной концентрации. При солюбилизации же независимо от используемого детергента может произойти разбавление мембранных белков и их разъединение. По закону действующих масс это приведет к диссоциации комплексов, в которых взаимодействие между компонентами не очень сильное.
Для изучения стехиометрии субъединиц и их ассоциации в очищенном комплексе используется всего несколько методов:
1) химическое сшивание;
2) количественный анализ N-концевых аминокислот;
3) определение отношения массы субъединиц в ДСН-полиакриламидных гелях путем определения интенсивности окрашивания, с помощью радиоавтографии или иммуноблоттинга.
Каждый метод имеет свои ограничения, но все они использовались на практике.
3.7 Изучение трехмерной структуры с помощью рентгеновской дифракции и реконструкции изображения
3.7.1 Кристаллизация мембранных белков
Наиболее детальную структурную информацию об очищенных мембранных белках можно получить, исследуя методом рентгеновской дифракции трехмерные белковые кристаллы. К сожалению, оказалось, что интегральные мембранные белки очень трудно кристаллизовать. Будучи удалены из своего естественного липидного окружения, неполярные участки липидных молекул склонны агрегировать с образованием неупорядоченных форм, непригодных для кристаллографического анализа. Ясно, что необходимы специальные методы, позволяющие обойти эти трудности, и в этом был достигнут определенный прогресс. Михель обратил внимание, что мембранные белки образуют кристаллы двух типов (рис.2).
Кристаллы типа I напоминают стопки мембран. В них осуществляется латеральное взаимодействие между неполярными участками, а мембраноподобные слои связывают полярные участки белков. Подобные кристаллы были получены для нескольких белков, но ни в одном случае их нельзя было исследовать с помощью дифракции с высоким разрешением. Кристаллы типа II стабилизируются за счет контактирования полярных участков белковых молекул, а небольшие амфифильные соединения или детергенты в основном заполняют промежутки между ними. Заметим, что очень важными являются размер, заряд и другие свойства детергентов; если эти параметры неблагоприятны, то детергент может дестабилизировать кристаллическую структуру. Кристаллы типа II образуют белки фотосинтетического реакционного центра Rhodopseudomonas viridis
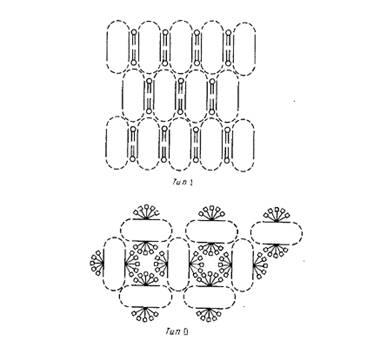
Рис.2. Два основных типа кристаллов мембранных белков.
Кристаллы типа I представляют собой двумерные стопки, упорядоченно расположенные в третьем измерении. В кристаллах типа II с гидрофобными поверхностями белков связаны молекулы детергента. Пунктиром отмечены гидрофильные домены белков.
Итак, мембранные белки можно кристаллизовать, и хотя число успешных попыток пока невелико, можно сделать несколько выводов, касающихся методологии кристаллизации.
1. Белки кристаллизуются вместе с детергентом.
2. Очень важен выбор детергента. По-видимому, наиболее пригодны цвиттерионные или неионные детергенты с высокой ККМ и малым размером мицелл
3. Кристаллизация облегчается в присутствии малых амфифильных органических молекул, по-видимому, влияющих на полярные концевые группы детергента.
4. Полиэтиленгликоль и сульфат аммония, обычно использующиеся при кристаллизации растворимых белков, применяются и для индукции кристаллизации мембранных белков.
3.7.2 Реконструкция изображения и двумерные кристаллы
Трехмерные кристаллы мембранных белков получить очень трудно, но многие из них образуют двумерные упорядоченные структуры. В некоторых случаях белки формируют такие структуры in vivo, (например, бактериородопсин в пурпурной мембране). При подходящих условиях такие белки, как и многие другие, образуют "двумерные кристаллы" при их очистке и реконструкции в присутствии фосфолипидов. Подобные двумерные упорядоченные структуры можно, использовать для получения трехмерной структурной информации с помощью электронной микроскопии и методов реконструкции изображения.
4. Пример структурных исследований мембранных белков.
4.1 Структура поринов
Порины - это основной класс белков, обнаруженных в наружной мембране кишечных бактерий
У Е. coli и Salmonella typhimurium выявлены три порина: OmpF, OmpC и PhoE. Эти белки имеют молекулярную массу примерно 35 000 и гомологичные аминокислотные последовательности. Порины экстрагируются из наружной мембраны с помощью ДСН в виде стабильных тримеров; их можно встроить в фосфолипидные бислои с образованием неспецифичных пор, способных пропускать малые (<600 Да) гидрофильные молекулы. По-видимому, именно они придают наружной мембране бактерий свойство молекулярного сита, позволяя питательным веществам проникать внутрь клетки, а отходам - выводиться наружу через неспецифические каналы.
Были проведены обширные структурные исследования белка OmpF, известного также под названием "матриксный порин"; в настоящее время осуществляется кристаллографический анализ, который позволит получить его структуру с высоким разрешением. Судя по данным о первичной последовательности, в молекуле нет никаких длинных гидрофобных участков, которые можно было бы идентифицировать как трансмембранные, и в среднем в ней содержится больше полярных аминокислот, чем неполярных. Однако трехмерная электронно-микроскопическая реконструкция изображения с использованием кристаллических пластинок реконструированного порина показала, что белок пронизывает бислой, причем за пределы мембраны выходят лишь небольшие участки молекулы. Методом негативного контрастирования были выявлены каналы, образуемые тримерами. Отдельные молекулы порина образуют у внутренней поверхности каналы, которые в середине бислоя сливаются в одиночный канал, открывающийся наружу. Есть и другие данные, свидетельствующие о том, что порин, несмотря на отсутствие в его молекуле гидрофобных участков, является трансмембранным белком. Иногда этот белок выполняет роль рецептора для бактериофага (для этого отдельные его участки должны быть экспонированы на наружной поверхности), а также проявляет сродство к компонентам клеточной стенки на периплазматической стороне. Очищенный порин можно встраивать в фосфолипидные бислои с образованием потенциалчувствительных каналов.
Данные инфракрасной спектроскопии, кругового дихроизма и широкоугольной диффузионной рентгеновской дифракции свидетельствуют о том, что две трети длины молекулы образует β-слой, а на долю α - спиралей приходится небольшая часть длины молекулы. Кроме того, эти исследования показывают, что β - цепи антипараллельны, ориентированы перпендикулярно плоскости мембраны не имеют среднюю длину 10-12 остатков, которых достаточно для пересечения неполярной области мембраны. Способ укладки β-цепей можно установить лишь с помощью рентгеновской дифракции. Как показывают модельные исследования, β-цепи могут быть уложены так, что образуется β-цилиндр, при этом полярные и заряженные аминокислотные остатки выстилают стенки наполненных водой каналов.
Все известные о структуре порина данные показывают, что гидрофобная
α-спираль не является его необходимым трансмембранным элементом. Это означает, что наиболее распространенные способы предсказания структуры трансмембранных белков имеют свои ограничения, поскольку они основываются на предположении, что пересечь бислой могут только гидрофобные сегменты. Точная структура порина до сих пор неизвестна; неясно также, сходна ли она со структурой других мембранных белков. Впрочем, имеются и другие белки наружной мембраны бактерий, которые характеризуются высоким содержанием β - структур.
Предположение о том, что необычная структура поринов связана с уникальной структурой и составом наружной мембраны бактерий, выглядит правдоподобно. Возможно, однако, что она обусловлена уникальностью способа образования больших водных каналов через бислой.
Заключение
Исследование мембранных белков все еще остается трудной задачей, требующей оригинальных подходов и нетривиальных решений. Тем не менее, каждый год появляются все новые сообщения о совершенствовании уже имеющихся методов. С разработкой новых методов очистки мембранных белков, основанных на применении различных детергентов, а так же с использованием методов секвенирования ДНК удается получить новую информацию о структуре мембранных белков. Данные по вторичной и четвертичной структуре очищенных мембранных белков можно получить с помощью биохимических и спектроскопических методов. Однако для построения моделей с высоким разрешением следует применять рентгеноструктурный анализ. Показательным в этом отношении является успех, достигнутый при изучении реакционных центров бактерий.
Список литературы
1. Геннис Р. Биомембраны: Молекулярная структура и функции: Пер. с англ. – М.: Мир, 1997,-624с.
2. Финдель Дж. Б., Эванз У.Г. Биологические мембраны. Методы: Пер. с англ. – М.: Мир, 1990, - 424с.
3. Фаллер Дж. М., Шилдс Д. Молекулярная биология клетки. Руководство для врачей: пер. с англ. – М.: Бином - Пресс, 2004,-272с.
4. Овчинников Ю.А. Биоорганическая химия Москва, Просвещение 1987.
5. Лефковитс И., Пернис Б. Иммунологические методы исследований(3 том) Пер. с англ. – М.: Мир, 1988,-530с.
6. Березов Т.Т., Коровкин Б.Ф. Биологическая химия. Москва, "Медицина", 1998 год,-704с.
7. Сим Э. Биохимия мембран: Пер. с англ. – М.: Мир, 1985,-110с.