Біологічний контроль збудників хвороб рослин
ЗмістВСТУП
РОЗДІЛ 1. Сучасний стан епідеміології вірусів вищих рослин
РОЗДІЛ 2. Шляхи розповсюдження вірусів рослин в природі та роль факторів навколишнього середовища
2.1 Клімат та погодні умови
2.2 Розповсюдженість вірусів. Біотичні фактори
2.3 Розповсюдженість вірусів. Абіотичні фактори
2.4 Шляхи передачі вірусів рослин в природних умовах
Розділ 3. Найбільш поширені вірусні захворювання серед вищих рослин
3.1 Кількісна характеристика та класифікація вірусів рослин
3.2 Віруси, які вражають певні види рослин
3.3 Віруси, які уражують широке коло рослин господарів
3.4 Боротьба із вірусними хворобами рослин
РОЗДІЛ 4. Біологічний метод: його суть та використання біопрепаратів
4.1 Біологічний метод
4.2 Ефективне застосування біопрепаратів при вирощуванні овочевих культур
4.3 Бактеріальні препарати і біогумат та їхні властивості
ПРАКТИЧНА ЧАСТИНА
Аналіз способів застосування бактеріальних препаратів і регулятора росту біогумату на овочевих культурах
Ефективність використання бактеріальних препаратів і регулятора росту біогумату при вирощуванні овочевих культур
Застосування бактеріальних препаратів і регулятора росту біогумату на помідорах
ВИСНОВКИ
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
Вступ
Актуальність. Протягом останнього часу в фітовірусологи, як і в інших молекулярно-біологічних дисциплінах, спостерігається значний інтерес до розвитку досліджень на популяційному та еколого-популяційному рівні, і ця тенденція є загальною для багатьох традиційних молекулярно-біологічних досліджень. Значні успіхи у фітовірусологи на молекулярно-біологічному рівні, такі як розшифровка структури генів, білок-білкової та білок-НК взаємодії, розшифровка АК-послідовностей та тривимірної будови структурних та регуляторних вірусних білків та багато іншого контрастує з нашими знаннями відносно поведінки вірусів рослин в природніх умовах біоцеозів. Поява в останні десятиріччя нового покоління діагностичних методів для визначення наявності фітовірусів (таких як ІФА, RIA, RIPA, ПЛР, ЛЛР, та ін) дали змогу на новому рівні вивчати розповсюдженість фітовірусів в навколишньому середовищі. Актуальність даного питання полягає в його як фундаментальності, так і в практичному значенні. Визначення ареалу розповсюдженості, механізмів передачі, кола рослин-господарів, реакції на зміни оточуючого середовища дає можливість не тільки більш повно характеризувати того чи іншого представника царства Vira, але і дасть змогу прогнозувати появу та розвиток вірусних хвороб і правильно виробляти стратегію і тактику боротьби з ними — запровадження стійких сортів, цілеспрямована боротьба з резерватами та переносниками, отримання безвірусного посадкового матеріалу.
Перспективним напрямком є моделювання розвитку хвороб, однак для вірусних хвороб рослин адекватні моделі практично відсутні, на відміну від фітопатогенних грибів та мікроорганізмів.
Встановлення можливих взаємозв'язків між наявністю вірусних антигенів в рослинах та ґрунті є дуже важливим у розумінні розповсюдження фітовірусів рослин та циркуляції їх в агроценозах.
Предмет дослідження – збудники хвороб рослин.
Об`єкт дослідження – застосування біологічного контролю збудників хвороб рослин.
Мета роботи – з`ясувати шляхи проникнення вірусних інфекцій в рослинні організми
Основні завдання:
1) охарактеризувати сучасний стан епідеміології вірусних захворювань рослин на Україні;
2) розглянути шляхи розповсюдження вірусів рослин в природі та роль екофакторів у цьому процесі;
3) дати стислу характеристику найбільш поширених в Україні вірусів рослин;
4) розглянути основні заходи боротьби з вірусами рослин;
5) проаналізувати суть та перспективи застосування біологічного контролю збудників рослин.
Розділ 1. Сучасний стан епідеміології вірусів вищих рослин
На порозі двадцять першого століття епідемії хвороб рослин все-таки призводять до значних втрат у продовольстві, промисловості, лісових та сільськогосподарських культурах всюди у світі. Населення нашої планети продовжує збільшуватись і якщо ми хочемо мати достатньо продовольства, одягу та помешкань для поточного та майбутніх поколінь, важливим є зменшення або й зовсім усунення втрат від хвороб рослин.
Епідеміологія хвороб рослин являє собою вивчення просторових та часових змін, що відбуваються під час епідемій хвороб рослин, які викликані популяцією патогенів у популяції рослин. Епідемія трапляється, коли відбуваються зміни у інтенсивності захворюваності серед популяції господаря у часі та просторі. Патогеном можуть бути гриби, бактерії, мікоплазмоподібні організми (МПО), віруси, віроїди, нематоди або паразитичні рослини. Рослини можуть бути частиною будь-якої сільськогосподарської, садової, лісової, або природної екосистеми. Взаємодія патогена та рослини-господаря, що призводить до розвитку хвороби, може трапитися у будь-якому наземному, водному, або штучно створеному середовищі. Сума усіх патогенів, господарів та середовищних компонентів являє собою рослинну патосистему.
Наука, що вивчає епідемії хвороб рослин (тобто, епідеміологія) має як описову, так і кількісну фази. Спочатку ставиться питання, наприклад, де епідемії трапляються, коли можуть епідемії очікуватися, які фактори відіграють роль у розвитку хвороби. Коли знання патосистеми збільшується, починаються питання більш кількісної природи, наприклад, скільки інфекційних одиниць патогену потребується, щоб започаткувати епідемію, скільки хвороба триває, як швидко буде хвороба розвиватися, та як далеко може інфекційна одиниця патогену розповсюдитись. Щоб відповісти на ці питання, використовується експериментальна та аналітична техніка класичної фітопатології та техніки, розроблені у агрономії, ботаніці, хімії, екології, ентомології, генетиці, математиці, метеорології, фізиці та статистиці.
Віруси та мікоплазмоподібні організми (МПО) рідко є окремими, незалежними інфекційними одиницями в оточуючому середовищі. Навіть частки цих патогенів звичайно пов'язані з рослиною-господарем або з переносниками (комахами чи нематодами). Таким чином, визначення рівня інфекційних одиниць в першу чергу робиться шляхом оцінки титру патогена в рослинах-господарях, або шляхом моніторингу популяції відомих переносників та визначення частки популяції переносника, що несе патоген рослин. У випадку титру вірусу точну кількість його не вимірюють, тоді як комаха або нема-індії, кожна з яких є переносником патогену, насправді є інфекційною одиницею або пропагулою епідемічного розвитку.
Зібрані зразки з інфекційними збудниками вірусної природи можна ідентифікувати за допомогою електронного мікроскопу, візуального мікроскопічного спостереження або шляхом біологічного тестування. Біохімічні технології можуть бути корисними для вірусів та МПО. При виборі методу моніторингу потрібно приймати до уваги складність ідентифікації, необхідний час та вартість витратних матеріалів.
Концепція збудника не зовсім чітка для вірусів та МПО. Хоча існують окремі частки або одноклітинні організми, вони мало впливають на епідеміологію хвороб, спричинених вірусами або МПО. Для багатьох з цих хвороб патоген передається з рослини на рослину за допомогою діяльності комах. Деякі ґрунтові віруси переносять нематоди або гриби. Обчислення кількості інфекційних одиниць поза рослиною-господарем роблять за допомогою збирання зразків векторів. Обчислення вірусів та МПО в їхніх рослинах-господарях для епідеміологічних завдань дає кращі результати при оцінці інтенсивності хвороби.
Хоча рослинні патогени переносять різні ряди комах, найбільш численні вектори має ряд Homoptera. Родини цього ряду включають горбаток, білокрилок, кокцид та попелиць. Взагалі, комахи Homoptera переносять 241 вірус та 45 МПО. Як і у випадку з повітряними грибами, комах можливо обчислити за їхнім накопиченням (зіткненням) із стаціонарними пастками, або їх концентрації в повітрі за допомогою всмоктувальних пасток. Далі, можливо визначити кількість особин, що несуть патоген та здатні до його трансмісії.
Оцінка хвороб рослин є одним з найбільш важливих та часто найбільш складних завдань у епідеміології хвороб рослин. Оцінка хвороби на даний час є підвалиною статистичного аналізу даних, зусиль з моделювання та інтерпретації патосистеми. Похибка, введена під час оцінки хвороби, може бути дуже малою на початку, але збільшитись при подальших епідеміологічних аналізах. Отже, ступінь впевненості у висновках та спроможність розрізнювати справжні ефекти не буде більшим, ніж рівень надійності при оцінці хвороби.
Оцінка хвороби звичайно триває багато часу і тому відносно дорога. Кропітке планування — як, коли, де та ким — хворобу буде оцінено, дуже істотне. Планування вимагає ясної концепції мети специфічного дослідження та докладне знання патосистеми.
Оцінки хвороби можуть бути кількісними, якісними, або комбінацією цих двох типів. Кількість наявної хвороби можна назвати поширеністю хвороби. До інтенсивності хвороби входять розповсюдженість хвороби та суворість хвороби. Додатково, такі характеристики, як концентрація або титр пропагативних або інфекційних одиниць (віріонів) у тканинах, можуть віддзеркалити інтенсивність хвороби. Якісні оцінки хвороби можуть включати як ринкову якість (вміст протеїну, олії, або вміст поживних речовин у господарі або Його продукті) так і такі фактори, як поживність, смак та запах.
Поширеність хвороби — це число одиниць рослин, що візуально уражено, звичайно до загального числа одиниць, що оцінено. Якщо розглядати рослину в цілому як одиницю, тоді поширеність хвороби є пропорція або процент рослин із симптомами хвороби. Поширеність хвороби може також бути оцінена, використовуючи окремі гілки, листки або коріння як рослинну одиницю.
Поширеність та суворість як виміри оцінки вживають разом, і правомірність кожного виміру повинна бути встановлена для кожної окремої патосистеми. Поширеність часто найбільш легка та найбільш швидка міра хвороби. Число зів'ялих рослин томату, почорнілі колосся пшениці, корінь із пухлинами або листя з некрозами чи мозаїчними симптомами можуть бути пораховані досить точно та акуратно. Звичайно поширеність є відповідною мірою хвороби для патосистем, у котрих один некроз на одиницю рослини є фатальним, також як і у випадку з хворобами в'янення та багатьма системними вірусними хворобами. Поширеність симптоматичних листків aбo гілок може також бути мірою хвороби для комерційних плодових дерев, тіньових лісів, тощо. Оцінка або вимірювання суворості хвороби часто більш тяжке та відбирає більше часу. Також, точність та відтворюваність може бути меншою, ніж у випадку поширеності. Суворість, однак, може бути більш важливою та корисною мірою дослідження хвороби для багатьох патосистем, що мають відношення до управління хворобами, динаміки чисельності та оцінки втрат врожаїв.
У більшості випадків, суворість хвороби розглядають як частку розглянутих рослинних одиниціь. Свідчення з фізіології рослин та досліджень втрат врожаїв показують, що врожай рослин може бути визначений із загальної суми уражених областей або, більш точно, із загальної суми здорових областей. Це примушує епідеміологів винайти шляхи для оцінки ступеню абсолютної суворості хвороби, хоча необхідно оцінити втрати врожаю. Однак на сьогоднішній день можуть бути застосовані декілька різноманітних шляхів репрезентації інтенсивності.
Терміни та частоти оцінок хвороби визначаються пагосистемою та метою оцінки. Оцінки можуть ґрунтуватися на календарних строках, фізіологічно-середовищних чинниках, або в залежності від стадій росту. Календарна шкала є найбільш часто використаною шкалою часу у епідеміологічних дослідженнях, що часто проводять через 5-, 7-, 10-, або 30-денні інтервали. Календарні оцінки є зручними та дозволяють скласти регулярний розклад роботи. Такі оцінки також забезпечують регулярно розмежовані дані для аналізу прогресу хвороби, у якому час використаний як незалежна змінна.
Фізіологічно-середовищна шкала часу використовує середовищні властивості або комбінації властивостей, що є важливими для фізіологічного стану господаря або патогену. Цей часовий масштаб ґрунтується на концепції, що певне число середовищних одиниць повинне бути акумульовано для того, щоб ключова фізіологічна подія трапилася. Число днів з певною температурою, вищою за мінімальну — приклад цього типу шкали часу. Цей тип шкали вимагає безперервного моніторингу змінних середовища (або доступ до таких даних). Складання розкладу у такому випадку оцінки дещо більш важке, ніж з календарною шкалою, якщо бажані регулярні інтервали одиниць. Такими шкалами користувались відносно рідко в фітопатології; однак, приклади є, та й подальше дослідження переваг цієї шкали часу виправдане.
Розширення фізіологічно-середовищної шкали базується на стадіях росту господаря. В межах цієї пов'язаної із господарем шкали, рослина сама слугує як інтегратор всіх середовищних ефектів. Припускаючи, що стадії росту ґрунтуються на ключових подіях у життєвому циклі господаря, оцінки хвороби при цих ключових стадіях забезпечують інформацію, що може бути корисною для аналітичної або модельної мети, для оцінки втрат врожаю, або для оцінки матеріалу у програмах вирощування рослин, стійких до хвороб.
Частота оцінок може залежати від частоти подій, вимог щодо інформації про різні стадії епідемії, або економічних міркувань. На фізіологічно-середовищній або шкалі стадій росту, частота специфічних подій (наприклад, акумуляція температурних днів або перехід до наступної стадії росту) визначить частоту оцінок хвороби. Якщо потрібен докладний статистичний або математичний аналіз епідемії, частота оцінки може визначатись потребою в специфічній кількості спостережень або потребі в спостереженнях під час "критичних" частин епідемії. Для епідемії, що триває 90 днів, тижневі оцінки дадуть 12-13 рівномірно розподілених спостережень. Ця кількість певно є достатньою для більшості випадків; однак, якщо економічні міркування диктують, що можна зробити менше ніж 12 спостережень, буде більш сприятливим зробити оцінки з нерегулярними інтервалами.
Економічні міркування при оцінці хвороби повинні бути врівноважені з потребою у специфічній інформації. Якщо час є обмежувальним фактором (як це і є звичайно), тоді необхідно визначити мінімальне число оцінок. Мета оцінки та знання джерел змін в межах та серед експериментальних одиниць буде диктувати, чи краще зробити більше спостережень у одному місці, або менше спостережень у багатьох місцях.
Можна зробити висновок про те, що маючи досить ґрунтовну теоретичну базу, ми ще не маємо більш-менш стандартних методик щодо об'єктивної оцінки розвитку вірусних інфекцій в агроценозах. Цей факт свідчить про актуальность цієї проблеми як її вузькому (вірусологічному) так і в широкому (сільськогосподарському та екологічному) розуміннях.
Розділ 2. Шляхи розповсюдження вірусів рослин в природі та роль факторів навколишнього середовища
2.1 Клімат та погодні умови
Віруси рослин мають здатність досить швидко розповсюджуватися в біоценозах. Вони можуть поширюватися завдяки переносникам (кліщі, попелиці, нематоди, гриби), передаватися при вегетативному розмноженні через бульби, живці, цибулини. Більшість фітовірусів передаються щепленим. Близько 90 різних вірусів рослин передаються насінням, деякі фітовіруси можуть викликати інфекцію без участі переносників, коли збудник знаходиться в грунті. Знаній екології збудника необхідне для використання необхідних засобів боротьбі проти нього (рис. 1.).
При ураженні рослин — господарів фітовірусами найчастіше розвивається системна реакція. При цьому інфекція часто протікає з яскраво вираженими симптомами у вигляді мозаїки, скручування, хлорозу, появи енацій та ін. Така інфекція може закінчуватись навіть загибеллю інфікованих частин рослин або і всієї рослини. Разом з тим, інколи перебіг інфекційного процесу проходить без чітких ознак хвороби, які помітні за габітусом рослин. Часто уражений організм може бути прихованим носієм інфекції. Продуктивність такі(Х рослин знижується в незначній мірі. Так реагують на вірусну інфекцію рослини-резерватори, переважна більшість яких відноситься до бур'янів.
Бур'яни (щириця біла, гірчиця біла, суріпиця звичайна, пирій повзучий та ін.) займають особливе місце серед факторів, що знижують врожай сільськогосподарських культур. Для цих рослий характерні велика енергія насіннєвого та вегетативної о розмноження, тривале зберігання схожості насіння. Вони сильно виснажують та висушують грунти, часто являються джерелом розповсюдження хвороб та шкідників сільськогосподарських культур. Джерелом вірусів служить також насіння бур'янів. Завдяки його достиганню та обсипанню до збору сільськогосподарських культур, насіння накопичується на полях у великій кількості. Так, в одному шарі товщиною 30 см знаходиться в середньому від 100 млн. до 4 млрд. і більше насіння бур'янів на 1 га. При обробці грунтів значна їх частина переміщується в верхній горизонт, і їх кількість тут в сотні і тисячі разів перевищує кількість насіння, що висівається. Крім того, грунт може бути забрудненшій кореневищами бур'янів, що також можуть бути уражені вірусами.
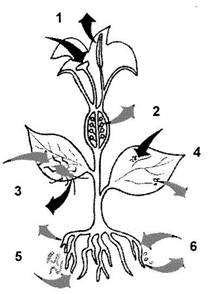
Рис. 1. Основні шляхи передачі фітовірусів.
1 - передача за допомогою пилку; 2 - передача за допомогою насіння; 3 - за допомогою векторів-комах; 4 - при прямому контакті з рослинами (механічному пошкодженні); 5 - за допомогою еекпюрів-нематод; 6 - за допомогою векторів-грибів.
Так, вірус мозаїки цукрового буряку уражує також щирицю, осот, кормові боби. Віруси, що викликають жовтяницю буряка, можуть перезимовувати в багатьох розповсюджених бур'янах.
Багато вірусів швидко мігрують завдяки переносникам. Переносниками можуть бути попелиці, цикадки, кліщі, нематоди, гриби та ін. Напевно, перше місце по передачі вірусів серед комах займають попелиці. Такі віруси, як вірус огіркової мозаїки, вірус жовтої карликовості ячменю, У-вирус картоплі, вірус мозаїки цукрового буряку, вірус жовтяниці цукрового буряку можуть передаватися різними видами попелиць. В деяких випадках, наприклад, між вірусом жовтої карликовості ячменю та попелицями, існує симбіотичний взаємозв'язок. При живленні попелиць на уражених цим вірусом рослинах підвищується їх плодовитість. Активними переносниками вірусу смугастої мозаїки пшениці є кліщі Aceria tulipae Keit та Асегіа tritici Schev. ВСМП може довгий час циркулювати в природі. Зимує вірус в інфікованих рослинах і кліщах. З весни та до збору врожаю кліщі інтенсивно розмножуються на рослинах, а потім мігрують на бур'яни (пирій повзучий, стоколос безостий та ін) і кукурудзу. Восени воші мігрують на сходи озимих та уражують їх вірусом (40).
Попелиці і кліщі, як і інші летючі комахи, є одними з важливих факторів, що визначають дальність розповсюдження віруса від джерела. Так, за один день попелиці відносяться вітром на десятки і навіть сотні кілометрів.
Грунт є також одним із джерел інфікування рослин вірусами. Потрапляючи в грунт, віруси здатні інфікувати нові рослини. Існує три групи вірусів, що передаються через грунт:
1. віруси, що передаються грибами;
2. віруси, що передаються нематодами;
3. віруси, переносник яких невідомий.
Крім вірусів, що переносяться або нематодами з родів Longidorus, Xiphinema і Trichodorus, або зооспорами ґрунтових грибів з родів Polymixa і Olpidium, існує група ґрунтових вірусів, представники якої "вільними" вібріонами зустрічаються в грунті і воді та можуть переходити без допомоги переносників від хворих до здорових рослин (non-vectored viruses), хоча деякі з них можуть переноситися нематодами або грибами.
Незважаючи на свою різноманітність, грунтові віруси мають ряд спільних ознак (за винятком ВОМ):
- не розповсюджуються літаючими переносниками;
- накопчуються в рослинах-господарях у високих концентраціях і виділяються через корені інфікованих рослин в грунт;
- надзвичайно стійкі поза живою клітиною;
- можуть інфікувати здорові рослини через корені без допомоги переносників;
- мають, як правило, широке коло рослин-господарів;
- більшість з них мають простий однокомпонентний геном;
- деякі з них можуть розповсюджуватись за допомогою безпосереднього контакту.
В грунт віруси рослин можуть потрапити як прямим (з листових, стеблових і кореневих залишків уражених рослин, та при віддачі вірусів коренями інфікованих рослин), так і непрямим шляхом (вірусовмісні екскременти та вимивання вірусів з контамінованих ґрунтів дощовими водами). Так, вірус огіркової мозаїки може проходити через травний канал людини або тварин, а потім зі стічними водами потрапляти в грунт і вражати рослини.
Джерелами контамінації "вільних" від вірусів ґрунтів або інших субстратів може бути, ймовірно, невелика кількість рослин, що постійно виділяють через корені вірус в навколишнє середовище. Деякі віруси настільки призвичилися до цього способу розповсюдження, що інфікують тільки кореневу систему рослин-господарів або окремі частини їх надземних органів. Діагностика в такому випадку ускладнена, і можна припустити, що віруси цієї групи розповсюджені більш широко, ніж думали до останнього часу.
На даний період в літературі описані 11 вірусів, у яких встановлена віддача вірусів через корені і 11 вірусів, у яких показана інфекція рослин через корені без переносників.
Численні віруси рослин містяться в ґрунті і в поверхневій воді, особливо в сільськогосподарських областях. Ступінь адсорбції і тривалість періоду, на протязі якого інфекційність зберігається і який може тривати тижні, а іноді і місяці, залежить від захисту вірусу і факторів зовнішнього середовища. Серед останніх це тип ґрунту, рівень вологості, рН, температура, вміст органічних субстанцій і звичайно зв'язана з цим біологічна активність середовища. Багато вірусів стабільні і здатні інфікувати рослини без допомоги векторів, а саме нематод та розповсюджених в ґрунті зооспор грибів. Передача вірусу з ґрунту та води ймовірно можлива через непошкоджені корені рослин. Віруси, які є в струмках, можуть переноситись на великі відстані. Іноді нові віруси, не знайдені у рослинах-господарях, були ізольовані з води та ґрунту.
До теперішньої о часу приблизно 9 вірусів були ідентифіковані і кілька інших ізольовані з ґрунту і води лісових екосистем. Вони головним чином належать до: Potex,- Tombus-, Tobamo-, Poty- та Necrovirus груп. Крім цього, ці віруси та принаймні 7 представників інших таксономічних груп, особливо Nepo- та Ilarviruses, були знайдені в деревовидних лісових рослинах. Такі збудники вірусних інфекцій можуть бути більш або менш шкідливі для інших видів рослин. Також багато вірусів було ідентифіковано в квіткових лісових рослинах. Ці патогени являють собою потенційну загрозу для економіки лісового господарства.
Невелика кількість досліджень поширення вірусів в цих субстратах (ґрунті та воді) спонукає нас проводити роботу по ідентифікації та розповсюдженості вірусних інфекцій в агроценозах України.
Останнім часом увага дослідників була привернута до вірусних систем збереження, як видно з екологічних робіт. Для деяких вірусів добре розвинена здатність до збереження на полі (дільниці) компенсується обмеженою здатністю до поширення на нові дільниці. Навпаки, інші віруси, особливо деякі з тих, що передаються персистентим способом повітряними векторами, широко передаються на нові дільниці, але не здатні до довгого збереження на даній дільниці (якщо мова йде виключно про агроценоз). Володіння ефективними способами як передачі, так і збереження, є характерною рисою деяких з більшості постійно циркулюючих вірусів.
Тому що віруси з однієї таксономічної групи мають подібну систему виживання, а різні типи рослинних угрупувань ймовірно мають схильність до різних видів систем виживання, можна очікувати, що специфічні групи вірусів будуть краще адаптовані до специфічних рослинних угрупувань. В середовищі, придатному для контактної передачі тобамо- та потексвірусів, рослини виробляють високу концентрацію відносно стабільних часточок в їхніх господарях. Інші адаптовані до культивованих рослин віруси (CULPAD viruses), включають іларвіруси, які головним чином можна знайти в дерев'янистих видах. Вони передаються пилком при запиленні рослин. Віруси, які переносяться за допомогою грибів (групи вірусу жовтої мозаїки ячменю), персистують на рестингспорах їхніх векторів і віддають перевагу монокультурам. З іншого боку, є віруси, адаптовані до диких рослин (WILPAD viruses) які включають багато груп, таких як непо-, тобра-, геміні-, лютеовіруси. Ці віруси мають широкий спектр господарів і довготривалу персистенцію на їхніх векторах. Ці властивості підходять до виживання в угрупуваннях, які складають багато видів рослин.
Екологія вірусів рослин, а віднині і епідеміологія вірусних хвороб рослин не повинна розглядатись як відбір незмінних зразків, вони можуть змінюватись під впливом багатьох факторів, таких як довгострокова зміна погоди, зміни в методах вирощування рослин, зміни екологічних параметрів ценозу. Для прикладу, вирощування рослин з використанням напівпрозорих пластикових листів в деяких країнах привело до поширення вірусів на захищених полях, а в Англії зростання заміщення ярого ячменя на озимий супроводжується зростанням кількості і поширенням вірусів, які є проблемними для цих культур.
Крім того, до цих "сторонніх" змін в методах культивування сільськогосподарських рослин є більш прямі наслідки від спроб контролювати віруси. Наприклад, вирощування культури томатів, що мають ген стійкості до ВТМ (Тm-1), призводять до селекції і поширення нестійких штамів, але ці штами, очевидно, мають лімітовану біологічну придатність, і коли тиск, нав'язаний стійкою культурою, знімається, вони зменшуються в поширенні.
Найімовірніше, віруси рослин протидіють стійкості, що контролюється одним чи двома генами господаря, більш повільно, або не контролюються взагалі. Це очевидно відбувається через те, що їхні геноми мають обмежену ємність кодів і їхній генний продукт є багатофункціональним, або через те, що їхні засоби передачі обмежують ступінь, до якого рослини з стійким генотипом піддаються інокулюму. В цій "боротьбі генів" нестійкі варіанти можуть з'являтися, але як вважається, можуть бути відносно невдалими. Протилежними в цій взаємодії є селективні ефекти дії вірусів на їхніх господарів, які звичайно проявляються після багатьох років. Таким чином, гени для толерантності або резистентності до інфекції звичайно знаходять на культивованих посівах на ділянках, де рослини постійно або регулярно піддаються вірусній інфекції довгий проміжок часу — резистентність кукурудзи до вірусу смугастості кукурудзи в Ісландії, резистентність картоплі до декількох вірусів в Південній Америці є тому прикладом.
В епідеміології географічний діапазон факторів, які працюють, є дуже важливим, тому шкала подій збільшується, що приводить до більшої складності системи. Є багато прикладів того, що можна домогтися при детальному вивченні розповсюдженості вірусів на специфічних ділянках. Важкі для розуміння з точки зору епідеміології вірусу великі континентальні земляні маси, де фактори, діючі на протязі сотень кілометрів, можуть привести до значних ефектів, і де щорічна сфера дії інфекції може піддаватися сильним флуктуаціям. В таких областях останні досягнення методів, заснованих на вивченні великомасштабних рухів повітряних мас, можуть частково принести успіх для картографії міграції повітряних векторів. Дійсно, такий спосіб вивчення впливу однієї частини континенту на іншій показаний на прикладі впливу великомасштабного переміщення вірусу коричневої плямистості рису.
Для аналізу причин і перебігів епідемій останнім часом з'явилося багато корисних методик. Велика кількість рослин може зараз бути протестована на присутність вірусної інфекції з допомогою чутливих серологічних методів, таких як ELISA. Досягнуто прогресу в детекції вірусів серологічними методами в індивідуальних векторних організмах, зібраних на полях, та різні генотипи вірусів можуть бути вивчені з допомогою гібрідізації комплементарної ДНК. Такою є робота по порівняльній екології різних штамів вірусів. Можна очікувати, що комп'ютерний аналіз може допомогти розібратися з великими масивами даних, одержаних після різнобічного вивчення і постійних записів стану зовнішнього середовища, а також допомогти зробити життєво необхідний висновок по поясненню подій та їх прогнозуванню.
Ситуація, що існує в дійсності по розвитку кожного вірусу в конкретній місцевості або в світовому масштабі, є результатом складної взаємодії багатьох фізичних та біологічних факторів. Розуміння екології вірусу щодо окремої культури та конкретної місцевості важливе для розробки відповідних методів боротьби із захворюваннями, що він викликає. Як і по відношенню до більшості інших облігатних паразитів, основними екологічними факторами, що слід розглядати, звичайно вважають шляхи передачі вірусів від рослини до рослини, а також шляхи, якими інші фактори впливають на розповсюдженість вірусу.
2.2 Розповсюдженість вірусів. Біотичні фактори
Вірус, що зберігає стабільність структури, знаходячись як всередині рослини, так і зовні і досягає високих концентрацій в тканинах, має, зрозуміло, більшу ймовірність вижити та розповсюдитись, ніж той, що цими властивостями не володіє. Наприклад, ВТМ може на протязі довгого періоду зберігатись в мертвому рослинному матеріалі, що знаходиться в грунті, котрий в такому разі стає джерелом інфекції для наступних культур.
Віруси або штами вірусів, що переміщуються по рослині повільно від того місця, де пройшло ураження, мають меншу ймовірність вижити і потім ефективно розповсюджуватись, ніж ті з них, що транспортуються швидко. Швидкість руху вірусів відіграє певну роль, якщо її розглядати з точки зору тривалості життя окремих рослин-господарів. Так, віруси, що уражують довгоживучі дерева та кущі, можуть рухатись по своїм рослинам-господарям значно повільніше, ніж ті, що інфікують однорічні рослини. Віруси, що можуть проникати в насіння і зберігатись там, мають суттєві переваги порівняно з іншими, з точки зору розповсюдження, а також здатності до виживання. Розповсюдження вірусу некрозу тютюну у більшості рослин-господарів обмежене кореневою системою, і існування цього вірусу в природі залежить від здатності грибів-переносників передавати його через грунт і уражувати таким чином інші рослини. Ймовірність виживання вірусу, що викликає швидко-прогресуюче системне захворювання, яке призводить до загибелі рослини, значно менша, ніж у тому випадку, коли вірус викликає у рослин слабкі або середньої тяжкості захворювання, при яких вони продовжують вегетувати та розмножуватись. Цикадки інфікуються спочатку штамами вірусу кучерявості верхівки цукрового буряку, що викликають слабкі симптоми на рослині-господарі. Вірулентні штами цього вірусу викликають загибель рослин до того, як на них закінчить, розвиток наступне покоління цикадок. Таким чином, в деяких умовах вірулентні штами мають тенденцію до самознищення. В країнах з високими температурами влітку може проходити інактивація вірусів in vivo в природних умовах. Так, в деяких районах Індії бульби картоплі, що зберігаються на звичайних складах, можуть звільнитись від вірусу згортання листків, в той час як бульби, що зберігаються в охолоджених сховищах, залишаються повністю інфекційними. Аналогічним чином рослини суниці звільнялись від вірусу зморшкуватості.
Таким чином основні фактори, що впливають на виживання штаму, такі:
- ефективна передача комахами або іншими засобами;
- більш швидке розмноження і більший врожай, ніж у конкуруючих штамів;
- розвиток слабкої або не надто сильної форми захворювання. Віруси широко варіюють по відношенню до спектру тих видів рослин-господарів, що вони уражують. Так, деякі віруси, що уражують полуниці, обмежені родом Fragaria. А вірус бронзовості томатів має дуже широке коло господарів, до якого входять як однодольні, так і дводольні. Відомо, що цей вірус уражає рослини 166 видів, що входять до 34 родин.
Віруси з дуже вузьким колом рослин-господарів, ймовірно, виживають тому, що ці рослини багаторічні, або розмножуються вегетативно, або внаслідок того, що такі віруси передаються насінням. Для багатьох нематод та вірусів, які вони переносять, характерний широкий спектр рослин-господарів, до яких входять і багаторічні деревоподібні рослини.
Оцінюючи віруси рослин в цілому, вважається, що найважливішим агентом розповсюдження, а таким чином, і виживання вірусів є літаючі комахи-переносники, що живляться соком рослин, зокрема попелиці. Шлях розповсюдження вірусу в межах окремої культури, а також швидкість та відстань, на яку його може бути перенесено, залежить від багатьох факторів (віруси, що передаються через повітряне середовище):
- джерело інокулюму; інокулюй може потрапити в культуру зовні, від хворих рослин в самій культурі, що мали вірус при передачі насінням або при вегетативному розмноженні, а також від
бур'янів та інших рослин, що знаходяться в цій культурі;
- кількість потенційно інфекційного інокулюму;
- природа та modus vivendi переносника (наприклад, щодо попелиць, то має значення чи це колонії, чи крилаті форми);
- передається вірус за допомогою стилету, є він циркулюючим чи розмножується в переноснику;
- час, коли переносник стає активним, відносно строків вегетації рослин даної культури;
- погодні умови.
Ван Дер Планк ще у 1946 році запропонував метод, за допомогою якого можна визначити, чи розповсюджується вірус в межах ділянки від ураженої рослини, чи він потрапляє зовні. Його засновано на припущенні, що вірус, який потрапляє зовні, буде викликати випадкове ураження рослин, тоді як при його розповсюдженні від хворої рослини до хворої, уражені рослини будуть розташовані групами. Таким чином, оцінюючи кількість пар уражених рослин відносно очікуваної, роблять висновок про походження вірусу.
Для вірусів, що розповсюджуються через грунт швидкість розповсюдження залежить від:
- типу переносника;
- структури, вологості, щільності і т.п. ґрунту;
- густини рослин-господарів;
- кліматичних умов;
- інших факторів.
Однак, відомі і виключення. Так, ВТМ — один з небагатьох вірусів, що в помітних кількостях поширюється через грунт без допомоги переносників.
Окрему групу феноменів складає перенесення вірусів на великі відстані. Більшість з таких явищ спонтанні і здебільшого не підлягають передбаченню. Вони порушують усталену рівновагу в ценозі, і тому довгострокові прогнози можливі лише на відносно стабільних системах, оскільки охоплюють коло тільки відомих характеристик.
На ураженість деякої культури захворюванням вірусної етіології значний вплив мають методи її вирощування та заходи обробки, що застосовують протягом багатьох сезонів. Тому будь-який прогноз має містити рекомендації щодо вдалого сівообігу, засобів обробки, термінів висіву та збору рослин. Вплив різного роду факторів, що мають значення у данному випадку, можна розглянути на таких прикладах.
Якщо поряд із полем озимини знаходиться поле ярої пшениці, ураженої вірусом смугастої мозаїки, то висів озимини до збору ярової пшениці призводить до великого ступеня її ураження ВСМП, який переноситься кліщами.
Тип сівообігу має деякий вплив на розповсюдженість вірусів, що здатні зимувати в бур'янах або у падалиці попередньої культури. В деяких культурах велика кількість спонтанно ростучих рослин, що містять вірус, може вегетувати довгий термін. Донкастер та Грегорі довели, що для того, щоб позбавитись від таких рослин картоплі, необхідно 5-6 років. А у випадку багаторічних культур кількість хворих р