Биологические особенности европейской ряпушки, необходимые для ее искусственного разведения
Россия, обладая значительным морским побережьем, а также громадным водным потенциалом внутренних водоемов, всегда являлась ведущей рыбохозяйственной державой в мире. В современных условиях, когда уловы океанической рыбы и других морепродуктов сокращаются, а рыбные запасы внутренних водоемов находятся в критическом состоянии, поддерживаются в основном за счет искусственного воспроизводства, единственным надёжным источником увеличения объемов пищевой рыбопродукции является аквакультура.
По официальным данным ФАО, в 2002 г. общее мировой вылов всех водных организмов, включая аквакультуру, составил 146 млн. т, в том числе промысел - 94,6 млн. т и аквакультура - 51,4 млн.т. Если объем промысла сохраняется в последние годы практически на одном уровне, то, по сравнению с 1990 г., объем продукции аквакультуры увеличился на 35 млн.т. При этом следует особо отметить, что весь этот объем приходится на пищевую продукцию, реализуемую в основном в живом и охлажденном виде.
По оценкам ФАО, аквакультура - самая динамично развивающаяся отрасль производства продуктов питания.
Высокие репродуктивные возможности рыб, быстрый рост при невысоких кормовых затратах, наличие в хозяйствах достаточного маточного поголовья, производство рыбы в местах ее потребления и независимо от внешнеполитических ситуаций позволяет уже в течение ближайших лет резко увеличивать объемы её выращивания.
Все изложенное убедительно свидетельствует о достоинствах и несомненных преимуществах аквакультуры. Можно с уверенностью утверждать, что аквакультура в России, в частности рыбоводство, имеет не только давние и глубокие корни, но и большие перспективы развития. Аквакультура имеет два основных направления. Первое - товарная аквакультура, у нас это товарное пресноводное рыбоводство. И второе - искусственное воспроизводство ценных видов рыб.
Учитывая, что объемы продукции морской аквакультуры у нас еще крайне незначительны, хотя перспективы огромные, основное внимание уделяют пресноводному рыбоводству и искусственному воспроизводству.
После распада СССР Россия оказалась самым крупным производителем продукции товарной аквакультуры. Однако и российские предприятия не миновал спад производства.
Однако в 1997 с. этот спад удалось остановить, и наметились определенные тенденции для роста производства товарной рыбы. В 2002г. хозяйствах было выращено более101 тыс. т. товарной рыбы и 600 млн. шт. посадочного материала различных видов рыб (Привезенцев,1991).
Помимо товарного рыбоводства важнейшее значение в отечественной аквакультуре имеет искусственное воспроизводство гидробионтов. Именно за счет искусственного воспроизводства во всем мире обеспечивается выращивание многих миллионов тонн морской, пресноводной рыбы и беспозвоночных. Россия обладает отлаженной системой воспроизводства рыбных запасов. В стране насчитывается более 150 рыбоводных предприятий, занимающихся получением и выпуском молоди осетровых, лососевых, сиговых, частиковых и других видов рыб. Основную роль в этой работе выполняют рыборазводные предприятия и организации Росрыбхоза и Госкомрыболовства России, ежегодного выпускающие во внутренние водоемы страны 7-8 млрд. экз. молоди и личинок ценных промысловых рыб.
Громадная площадь естественных водоемов, водохранилищ, водоемов комплексного назначения, значительный прудовый фонд, большое число хозяйств индустриального типа, отработанные технологии культивирования гидробионтов, наличие квалифицированных специалистов - все это является основой для увеличения продукции в России. Развитие аквакультуры должно постоянно сопровождаться проведением прикладных исследований в области кормления и технологий выращивания рыб, генетики и селекции, поддерживания нормального физиологического состояния культивируемых объектов.
Знание биологии закономерностей воспроизводства рыб в равной степени необходимо как при разработке мероприятий по охране и рациональной эксплуатации, так и при подборе базовых для рыбоводства популяций и стад. Естественно, что и выбор водоемов для интродукции и акклиматизации новых видов, для организации интенсивных форм хозяйства (например, товарного рыбоводства) нельзя сделать без глубокого знания биологии разводимых видов (Мухачев,2005).
Поэтому целью данной работы является описать биологические особенности европейской ряпушки в связи с его искусственным разведением. Поставленная цель определила следующие задачи:
- дать биологическую характеристику европейской ряпушки в связи со средой обитания и образом жизни;
- определить влияние различных факторов среды на европейскую ряпушку;
- изучить различные методы управления половыми циклами у европейской ряпушки;
- рассмотреть биологические основы кормления европейской ряпушки;
- описать способы транспортировки икры, личинок, молоди и взрослых особей европейской ряпушки;
- охарактеризовать биологические основы акклиматизации.
Глава 1. Биологическая характеристика европейской ряпушки в связи со средой обитания и образом жизни
1.1 Общая биология вида
Тип: Хордовые (Chordata)
Под тип: Черепные (Craniata)
Над класс: Челюсноротые (Gnathostomata)
Класс: Рыбы (Pisces)
Над отряд: Костистые рыбы (Teleostei)
Отряд: Лососеобразные (Salmoniformes)
Под отряд: Лососевидные (Salmonоidei)
Семейство: Сиговые (Coregonidae)
Род: Сиги (Coregonus)
Вид: Европейская ряпушка (Coregonus albula L)
Европейская ряпушка (рис. 1) относится к семейству лососевых (Salmonidae), роду сигов (Coregonus). Характернейшей особенностью всех видов ряпушек, отличающей их от остальных видов рода Coregonus, является верхний рот. Этим обусловлены занимаемые ряпушкой экологические ниши и характер ее питания. Европейская ряпушка – типичный лимнофил, и вполне правомерно назвать ее озерной (Потапова,1978).
Как у всех сиговых рыб, тело серебристо-белого цвета, нет яркой окраски, как у лососевых, по форме больше похоже на тело карповых, но имеется жировой плавник. Один спинной и один анальный, все плавники без колючих лучей. Тело покрыто крупной циклоидной чешуей. Три последних хвостовых позвонка загнуты вверх, преуральный и уральный позвонки не сливаются в один конечный. У молоди, как правило, не бывает поперечных темных полос. У ряпушки рот верхний, нижняя челюсть заметно выступает вперед и вверх, верхнечелюстная кость заходит за вертикаль переднего края глаза. Глаз большой. У европейской ряпушки в отличие от сибирской С.sardinella нежная и легко опадающая чешуя, антидорзальное расстояние (AD) больше 42% длины тела и меньше число позвонков (54-59, чаще 55-56).
D III-IV 7-11; A III-IV 9-14; P I 11-16; V II 8-11. Жаберных тычинок 35-58, чаще 40-44. число чешуй боковой линии 67-98. Пиларических придатков 40-88. Кариотип: 2n = 80, NF = 96. однако у европейской ряпушки отмечена изменчивость кариотипа, описаны добавочные B– хромосомы, а также наличие половых хромосом, поэтому кариотип самок – 2n = 80 + В при NF = 96 + В, а у самцов – 2n = 81 + В при NF = 97 + В (Решетников, 1995).
Ряпушка достигает половой зрелости в двухлетнем возрасте, длина тела обычно достигает 20-26 см, масса 25-50 грамм. Нерестится поздней осенью или в начале зимы в период ледостава на твердом каменисто-песчанном грунте при температуре несколько выше 00С. Плодовитость – около 3-5 тыс. икринок. Выклев личинок из икры проходит весной в марте-апреле (Анисимова, 1991).

Рис. 1. Европейская ряпушка.
Вид европейская ряпушка Coregonus albula обитает в озерах (озерных системах) преимущественно в бассейна Балтийского моря и частично в бассейнов Белого (Карелия, Кольский полуостров, Архангельская область, включая Соловецкие острова), Баренцево (Кольский полуостров) и Каспийского (верховье бассейна Волги) морей. Имеется в водоемах Ирландии и Великобритании. Населяет реки и озера от Дании, юга Норвегии и Швеции (западная граница распространения вида) до Печоры и Вашуткиных озер (Решетников, 1980). В России южная граница проходит по Псковско-Чудскому водоему, озерам Верхней Волги (Селигер, Вселуг, Белое, Переславское). Общее количество озер населенных естественными популяциями европейской ряпушки в ее исходном ареале, составляет около 1000 (Потапова, 1978).
Жизненный цикл ряпушки подразделяется на ряд этапов и периодов характеризующихся спецификой развития, особенностями взаимоотношения со средой и определенной смертностью.
Этап неполовозрелого организма делиться на мальковую стадию которая длится около 3 месяцев (1-4 месяца), и стадию полового созревания – около 12-13 месяцев (с октября до октября – ноября следующего года). Половой зрелости (первого нереста) ряпушка достигает на втором году жизни (в возрасте 16-17 месяцев). Этап половой зрелости характеризуется ежегодным нерестом. Об этапе старости в отношении ряпушки можно говорить весьма предположительно: старость, видимо, наступает после 5-6 нерестов, т.е. в возрасте 7-8 лет.
На этапе половой зрелости организм ряпушки ежегодно претерпевает сходные сезонные изменения физиологического состояния, отражающиеся в поведении и завершающиеся нерестом. Поэтому можно говорить о годовых циклах в онтогенезе рыбы и давать внутригодовую периодизацию их.
Годовые циклы рыб (как и других животных) являются отрезком онтогенеза, на протяжении которого осуществляется вся совокупность жизненных процессов, обеспечивающих нормальное существование популяции. Годовые циклы делятся на периоды. Периодами годового цикла называют качественно своеобразные устойчивые состояния, через которые на протяжении года проходит популяция животных. Из внешних факторов особенно большое влияние оказывают сезонные ритмы температурного, газового и солевого режимов, обеспеченность пищей и т.д. В связи с этим основные процессы жизнедеятельности рыб – созревание и нерест, рост, накопление энергетических резервов – оказались приуроченными к наиболее благоприятным сезонам года.
На основании изучения физиологического и биохимического состояний ряпушки можно выделить 5 периодов: нерестовый, посленерестовый (предзимовальный), зимовальный, нагульный и преднерестовый.
Продолжительность нерестового периода различна от 3 недель до 1,5 месяца. Обычные сроки нереста: октябрь, реже ноябрь, декабрь, январь и очень редко март-апрель (озера Финляндии). Самки выметывают всю икру сразу. Созревание самцов более растянутое; сперма выделяется порциями. После нереста вес тела и тушки самцов и самок минимальны. Это объясняется как выметом половых продуктов, так и расходованием резервных веществ. Поступление энергии с пищей ограничено, так как нерестующая ряпушка не питается.
Нерестовый период характеризуется отрицательным энергетическим балансом и снижением содержания общих липидов и липидных фракций во всех органах, за исключением нервной ткани.
Посленерестовый период, кратковременный (от 2-3 недель до 1 месяца), ряпушка держится на нерестилищах и в открытой части озер. Коэффициент зрелости, показатели упитанности самцов и самок минимальные в годовом цикле. Отличительной чертой этого периода является интенсивное питание. Это способствует восстановлению пластических веществ и обеспечивает подготовку к длительной зимовке.
Зимовальный период в годовом цикле ряпушек и других рыб умеренной зоны зимовка длится 6-7 месяцев ( с ноября до середины мая). Однако это не пассивный период. В первую половину зимовки (декабрь-февраль) питание ряпушки носит поддерживающий характер. Интенсивность питания низкая (около половины рыб с пустыми желудками). Весовой и линейный рост отсутствует. Ряпушка малоактивна, держится в открытых участках озера и предпочитает поверхностные горизонты – непосредственно подо льдом.
Во второй половине зимовки (с конца марта – начала апреля до таяния льда), когда начинается прогрев водных масс и развитие зоопланктона, пищевая активность ряпушки несколько повышается, и к моменту таяния льда (май–начало июня) у самок появляется полостной жир на внутренностях. Во время зимовки обменные процессы ряпушки резко понижены. Однако, не смотря на низкие температуры в подледный период и замедленный метаболизм, в организме идут процессы, связанные с овогенезов, и к началу нагула (май) ганады переходят из VI во II стадию зрелости.
Нагульный период начинается сразу после вскрытия водоемов и продолжается до середины или конца августа. Он характеризуется усиленным откормом ряпушки. В начале нагула, когда еще зоопланктон развит слабо, старшие возрастные группы ряпушек потребляют личинок хирономид, ручейников и поденок. К концу нагульного периода завершается линейный рост самцов и самок. Коэффициент жирности и упитанности достигает максимального значения.
Преднерестовый период (конец августа – первая половина октября). Этот период характеризуется изменением направленности жирового обмена – от накопления резервных веществ к их интенсивному расходованию. Продолжается питание ряпушки и в течение 1.5 – 2 месяцев резко возрастает масса гонад (особенно яичников). Гонады переходят из III в IV, а затем и в V стадию зрелости. Коэффициент зрелости крупных ряпушек 13 – 23% у самок и 1.6 – 2.6% у самцов.
Для ряпушек характерно сравнительно раннее созревание и единовременный нерест. Размерно-весовой состав впервые нерестующей ряпушки разных озер имеет существенные различия. О ходе созревания ряпушки можно судить по непосредственному наблюдению за развитием и изменением веса гонад. Уже в конце августа и начале сентября у сеголеток (возраст 4 – 5 месяцев) пол определяется невооруженным глазом. Гонады самцов и самок в октябре переходят во II стадию зрелости, которая продолжается до конца июля – начала августа следующего года. После первой зимовки, в мае – июне, у самцов коэффициент зрелости колеблется в пределах 0,04 – 0,63, а у самок – 0,11 – 1,28% (II стадия зрелости, 1 год). К концу августа он повышается до 1,5 – 1,6 у самцов и 1,1 – 5,4% у самок. К этому времени гонады достигают III стадии зрелости, в течение сентября и первой половины октября более интенсивно идут процессы ово- и сперматогенеза. У самцов в это время стадии зрелости гонады IV – V, а у самок IV (возраст 1+). Переход в V («текучую») стадию обычно происходит в течение нескольких дней перед нерестом. Икринки становятся прозрачными и свободно лежат в полости тела. Как правило, ряпушка икру выметает полностью, остается лишь темно-бурая строма. У самцов семенники уменьшаются в размере и приобретают красноватую окраску (стадия зрелости VI – II). Вес гонад зимой почти не увеличивается и составляет у самок в среднем около 0,93% от веса тела, а самцов – 0,3 – 0,6% от веса тела. Созревание гонад для повторного нереста (трехлетки, четырехлетки и т.д.) имеет сходный характер.
Ряпушка относится к осенненерестующим рыбам. Сроки размножения определяются температурным режимом водоемов и изменяются в зависимости от гидрометеорологических условий года. Обычно ряпушка нерестится со второй половины октября и до декабря. Но в северных широтах (озера бассейна Ладожского озера, озера Финляндии) были обнаружены популяции европейской ряпушки, которые нерестятся в зимний период (декабре – январе). Нерестилища осеннеенерестующей ряпушки располагаются на глубинах до 10 м, а зимненерестующей – на глубине 10-20м. Зимненерестующая ряпушка отличается от осенненерестующей большим диаметром глаз и населяет более глубоководные участки. Представляет интерес находка весенненерестующей ряпушки в озерах Швеции. Она нерестует в марте – апреле на глубинах 25-28м. Морфометрических различий между осеннеенерестующей и зимненерестующей ряпушкой не обнаружено за исключением разного числа позвонков. Нерестилища ряпушки в большинстве водоемов располагаются на глубинах менее 10 м на твердых грунтах. Однако имеются сведения о нересте ряпушки на значительных глубинах (Потапова, 1978).
1.2 Эмбриональное развитие
Через 3-5 мин. после оплодотворения и пребывания в воде между яйцом и его оболочками проявляется узкая щель, начинается образование периветелинового пространства – «набухание» икры (рис. 2 а, б). Диаметр набухания икры достигает 1,5-2,2 мм. По мере роста большого периветелинового пространства в яйце заметно уплощается анимальная область. Одновременно с этим процессом происходит формирование бластодиска. Объем сформированного бластодиска равен 15-20% объема желточного мешка, диаметр его 1-1,2 мм (рис. 2 в, г). Жировые капли концентрируются у основания бластодиска, почти в одной плоскости, в то время как остальная часть желточного мешка их не содержит.
Образование периветелинового пространства и бластодиска происходит у оплодотворенных яиц одновременно, и отличить у их в этот период нельзя. Морфологически их различия проявляются после начала дробления оплодотворенной икры, и определить это можно только под микроскопом. У оплодотворенных яиц при содержании их во влажной атмосфере, на рамках изотермического ящика при среднесуточной температуре + 2,50С, уже 16 часов после оплодотворения борозда первого дробления, уже глубокой щелью разделяет светлую анимальную область, где происходит формирование зародыша (рис. 3 а, б). через 30 часов зародыш развивается до стадии 4 бластомеров (рис. 3 в).
Стадия 16 бластомеров наступает через 50 часов после оплодотворения (рис. 2 г), а через 96 часов при среднесуточной температуре + 2,50С, уже хорошо выражена стадия крупноклеточной и бластомерной бластулы с четко различимыми бластомерами. Через 6 суток инкубации при среднесуточной температуре + 1,80С зародыш развивается до стадии мелкоклеточной или «эпителиальной» бластулы (рис. 3 д, е).
Гаструляция зародыша начинается на 14-15 день инкубации (средняя температура 0,90С, сумма среднесуточных температур 14,40С) (рис. 4 а). Поверхность зародышевого диска уплощается и происходит обрастание желточного мешка бластодермы. Еще до полного обрастания и закрытия бластопора происходит формирование зародышевой пластинки. Через 9 дней после начала гаструляции зародышевая пластинка уже хорошо сформирована (длина 1,5 мм) и четко видна дифференцирующая нервная трубка и хорда (рис. 4 б). к этому времени бластодерма обрастает в желточный мешок на 80-85% (диаметр бластопора 1,5 мм) (рис. 4 в, г). Закрывается бластопор на 27-28 сутки развития (средняя температура 0,70С, сумма среднесуточных температур 190С) (рис. 3 в). К этому времени уже дифференцировалась головная пластинка. Происходит формирование зачатков глазных пузырей и отделов головного мозга.
У 42-дневного зародыша (средняя температура 0,60С, сумма среднесуточных температур 23,80С) начинается рост хвостовой почки. В туловище 18-20 пар мезенхимных сегментов. Хорошо развиты глазные пузыри. Образуются зачатки хрусталика глаза. Становятся видны обонятельные плакоиды, дифференцированы зачатки переднего, среднего, заднего отделов спинного мозга. Начинается обособление мезенхимы в области жаберных дуг. Длина эмбриона 3,2 мм. В процессе роста «хвостовой» почки возникают новые миотомы и соответствующие участки нервной системы и хорды (рис. 4 д).
На 60 день развития эмбриона (сумма среднесуточных температур 31,20С) хорошо видны зачатки плавников. В это же время на наружных мешках глазных бокалов появляются первые признаки черного пигмента. Начинается дифференцировка жаберных крышек. Образуется зачаток мозговой воронки и разрастаются отделы среднего мозга и мозжечка. Количество миотомов 30-32. Хвостовой и головной отделы приподнимаются над желточным мешком. Зародыш достигает 4,3 мм и составляет ¾ желточного мешка. Жировых капель – 2, каждый диаметром – 0,5 мм, они располагаются у головного отдела зародыша и имеют несколько удлиненную форму.
На 72 день развития (средняя температура – 0,60С) возникает плавниковая кайма – сначала вокруг хвостового отдела, а в дальнейшем вокруг всего тела эмбриона. Грудные плавники еще не имеют формы складки. Хорошо заметен зачаток печени, большой задний отдел кишечника. Происходит быстрое накопление пигмента на поверхности зачатка ретины и глаза делаются видны невооруженным глазом (рис. 4 е). пигментация туловища зародыша и желточного мешка начинается на 106 день развития (средняя температура 0,50С) при длине 6,2 мм. Крупные пигменты клетки звездчатой формы – меленофоры – в начале единично появляются в передней половине туловища и на ближайших тела участках желточного мешка. В дальнейшем их количество увеличивается и они располагаются вдоль всего тела в 2 ряда по верхнему и нижнему краю сегментов. В тоже время начинается образование зачатка хвостового плавника (происходит скопление мезенхимы в нижней части плавниковой складки). Образование зачатка спинного плавника происходит через 132 дня после оплодотворения икры (средняя температура 10С, сумма температур 142 0С). В это же время начинается пигментация головы и зародыша. Первая и вторая большие клетки появляются на затылочной части черепа. Число миотомов – 58-60. Во всех отделах кишечника есть полость. Хорошо видны складки в зачатке желудка. Сердце пульсирует и можно наблюдать слабый прерывающий ток крови. К этому времени на голове и передней части желточного мешка сформированы железы вылупления. Непосредственно перед вылуплением у зародыша 60 сегментов. Диаметр желточного мешка 1,1-1,2 мм, т.е. за весь период эмбрионального развития его диаметр уменьшился примерно на 0,5 мм. На жаберных дужках начинают дифференцироваться жаберные мешки. При средней температуре за весь период инкубации 1,20С (сумма среднесуточных температур 165,40С). Массовое вылупление личинок происходит через 137 дней после оплодотворения икры (рис. 3 е).
Средняя длина выклюнувшихся личинок равна 9,5 мм, окраска их сероватая, покров прозрачный. Личинки сохраняют зародышевую пигментацию. Тело окружено сплошной плавниковой каймой, которая прерывается только в области ануса. Зачаток спинного плавника представлен хорошо выраженной мускульной почкой и скоплением мезенхимы. Грудные плавники хорошо развиты, их лопасти имеют округлую форму. Зачаток брюшного и жирового плавников еще не различим. Хорошо развиты и функционируют кровеносная система желточного мешка. Желточный мешок удлиненной, овальной формы, вытянутый вдоль тела личинок, в плавниковой кайме появляются выемки между будущим спинным, жировым и хвостовым плавниками.
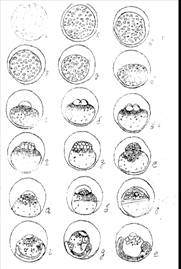
Рис. 2, 3, 4. Эмбриональное развитие
Через 6-7 дней после вылупления личинки переходят на активное питание. Желточный мешок весь почти резорбируется, а жировая капля еще сохраняется. Жаберные лепестки развиты хорошо. Усиливается пигментация личинок. На голове и спинной стороне тела развит пигмент. Черные пигментные клетки довольно густой сетью покрывают затылочную часть головы. Меланофоры появляются в хвостовом отделе под хордой и называются спинной нервной трубкой. Длина личинок в среднем 10,2 мм.
1.3 Постэмбриональное развитие
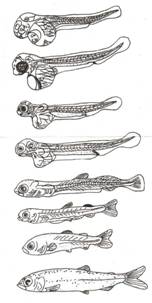
1 – этап желточного питания
2 – этап смешенного питания
3 – этап питания микропланктоном
4 – этап питания мезопланктоном.
Мелкий зоопланктон не едят, а употребляют личинки тендипендии. Темп роста резко возрастает. Этап завершается при длине около 5 см, после чего наступает мальковый период развития. К этому времени тело покрывается чешуей, остаток плавниковой каймы полностью исчезает. Длительность личиночного развития 30-36 суток (рис. 5 е, ж, з) (Пономарева, 1996).
а
б
в
г
д
е
ж
з
Рис. 5. Постэмбриональное развитие.
Европейская ряпушка обитает в озёрах преимущественно в бассейнах Балтийского моря и частично в бассейнах Белого, Баренцева и Каспийского морей. Половая зрелость наступает на втором году жизни при длине тела 20 – 26 см., и массе 25 – 50 г. Нерест поздней осенью в начале зимы при температуре несколько выше 0 0С. Плодовитость 3 – 5 тыс. шт. икринок. Промысловый вид, но численность не велика.
Глава 2. Влияние различных факторов среды на европейскую ряпушку
Рыбы, как и все другие организмы, обитающие в водоемах, находятся в тесном взаимодействии с абиотическими факторами среды – совокупность условий неживой природы, и биотическими факторами среды, включающие в себя разнообразные отношения между организмами. Грунт и вода с ее химическим составом и физическими показателями являются абиотическими факторами среды, действующими в водоемах.
Внешняя среда влияет на все жизненные процессы, происходящие в организме рыбы: дыхание, питание, кроветворение и кровообращение, нервную деятельность, размножение, рост и развитие. Рыба на разных стадиях своего развития и в различные периоды жизни не одинаково реагирует на условия внешней среды.
Богатство и многообразие ихтиофауны водоемов зависит от многих факторов среды, действующих на организм рыбы, в частности от площади и глубины водоема, характера его грунта, от населяющих его бактерий, растений и животных, от гидрологического и гидрохимического режима и т.д. чтобы правильно оценить ту огромную роль, которую играют факторы внешней среды в жизни рыб, достаточно рассмотреть лишь наиболее важные из них. С которыми чаще всего приходится сталкиваться специалисту, работающему в области рыбоводства: температуру воды, освещенность, уровень и течение воды, гидрохимический режим и кормовую базу (Иванов, 1988).
2.1 Влияние температуры
Температура воды является одним из факторов, оказывающих большое воздействие на отправление жизненных функций рыбы, определяющих ее рост и развитие. Этот фактор действует на европейскую ряпушку как непосредственно – изменяя интенсивность ферментативных процессов, происходящих в организме, активность потребления пищи, характер обмена веществ, ход развития половых желез и прочее, так и косвенно, оказывая свое влияние на улучшение или ухудшение развития естественной кормовой базы.
Температура является также внешним стимулом, определяющим для физиологически подготовленного организма рыб начало миграций, нереста и зимовки.
Температурные условия, при которых все жизненные процессы протекают в организме нормально принято называть оптимальными. Исходя из оптимальных температурных условий, все виды рыб условно подразделяют на теплолюбивых и холодолюбивых.
Ряпушка относится к холодолюбивым рыбам, которые нерестятся в основном осенью при температуре воды не выше 10-140С. Икра этих рыб развивается при температуре воды от 0 до 140С. Холодолюбивые рыбы наиболее интенсивно питаются и растут при температуре воды 8-150С. При дальнейшем повышении температуры воды у этих рыб резко снижается двигательная активность, интенсивность питания и замедляется рост. Резкое изменение температуры воды, если даже она и не выходит за границы оптимальных температурных условий, вызывает у европейской ряпушки нервный шок, который приводит обычно к гибели (Иванов, 1988).
Температурные условия водоема оказывают влияние на ряпушку как косвенно – через обеспеченность пищи (качественное и количественное развитие рачкового планктона), так и непосредственно, изменяя уровень обменных процессов в организме рыбы. Оптимальными условиями для питания ряпушки является температура 15-160С, а при повышении ее до 18-200С интенсивность потребления пищи ряпушкой снижается (Покровский, 1961).
В глубоководных озерах условия нагула ряпушки более благоприятны в теплые годы, так как она может использовать кормовые ресурсы придонных слоев воды. В мелководных озерах, прогревающихся летом до 18-200С, обитает европейская ряпушка, которая приспособилась к более высоким температурам. В ряпушковых озерах осенняя гомотермия в зависимости от гидрометеорологических условий года наступает в период с конца сентября до начала ноября. Это, вероятно, обусловливает растянутость сроков нереста ряпушки. При температурах близких к 2-40С наблюдается зимний нерест. Зимой температура придонных слоев воды обычно выше температуры поверхностных (около 40С). В малых и мелководных озерах, в первую половину зимы температура придонных слоев воды ниже 2-30С и только к концу марта повышается до 40С. В глубоководных озерах в течение зимы температура воды остается низкой 0,2-0,80С в поверхностных слоях, 1,10С на глубине 4 м, 1,9 и 2,30С на глубинах 20 и 27 м, а на глубине 30-32 м 1,3-1,70С.
Таким образом, озера, населенные европейской ряпушкой относятся к эпитермическому и метатермическому типам. Эпитермические озера умеренной зоны характеризуются устойчивой гомотермией в течение всего периода открытой воды.
Температура воды оказывает также большое влияние на развитие в водоеме кормовых организмов, являющихся пищей для ряпушки. Отклонения температур воды вызывают изменение численности и биомассы зоопланктона и бентоса, а это в свою очередь влияет на динамику численности рыб.
Интенсивность питания европейской ряпушки в значительной степени зависит от температуры. Повышение температуры вызывает ускорение переваривания пищи. В связи с этим повышается интенсивность приема пищи при более высоких температурах. Для каждого вида существуют температурные пределы, за которыми питание прекращается, и оптимальные температуры, при которых рыбы наиболее усиленно питаются. Так, для холодолюбивой европейской ряпушки оптимум питания приходится на 12-140С. А в жаркие летние дни, когда вода у поверхности прогревается до 200С, пищевая активность ряпушки снижается, и она уходит в придонные слои (Потапова, 1978).
Исходя из изложенного выше, можно сказать, что температура воды имеет огромное значение в жизни европейской ряпушки.
2.2 Влияние освещенности, уровня и течения воды на европейскуюряпушку
Освещенность, уровень и течение воды, как и температура, играют большую роль. Известно, что в результате таяния снегов и дождей повышается уровень воды в реке, увеличивается скорость течения, мутность и, следовательно, снижается степень прозрачности воды, которая существенно влияет на ее освещенность.
Прозрачность воды зависит от содержания в толще воды неорганических и органических взвешенных частиц, а также мельчайших растительных и животных организмов. Чем больше в воде содержится взвешенных частиц и микроорганизмов, тем меньше ее прозрачность и наоборот. Поздней осенью и зимой вода бывает более прозрачна, чем весной и летом, так как весенний паводок и летние ливневые дожди приносят в водоем большое количество неорганических и органических веществ, в это же время года в воде бурно развиваются одноклеточные водоросли и беспозвоночные животные. От освещенности у большинства рыб зависит строение органа зрения, которое играет значительную роль при ориентировке во время движения и суточном ритме активности. Освещенность влияет на развитие рыб. Так у многих видов ряпушек в эмбриональный период нарушается обмен веществ, если развитие происходит в несвойственных для них условиях освещенности (Иванов, 1988).
Освещенность влияет на строение органов зрения. Европейская ряпушка является планктофагом, а для них характерны сравнительно крупные глаза. По этому признаку выделяют популяции ряпушек обитающих в водоемах с разной прозрачностью. Так, риндозерская, топозерская и вендюрская ряпушка имеет крупные глаза в связи с обитанием в водоемах с низкой прозрачностью воды. А популяция европейской ряпушки с мелководного озера Урос характеризуется значительно меньшим диаметром глаза, так как обитает в водах с более высокой прозрачностью (Потапова, 1978).
2.3 Влияние гидрохимических показателей на европейскую ряпушку
Гидрохимический режим водоема оказывает на европейскую ряпушку такое же сильное воздействие, как все перечисленные выше факторы среды. Он зависит от химического состава воды, ее способности растворять жидкие, твердые и газообразные вещества. Совокупность указанных веществ, их характер и количество во многом предопределяют условия жизни рыб в водоемах.
Солевой состав воды. Минеральные соли растворенные в воде, и соленость в различных водоемах неодинакова. Морская вода резко отличается от пресной по своему солевому составу. В морской воде преимущественно растворены хлористые соли, а в пресной преобладают углекислые и сернокислые соли, обуславливающие соответственно «жесткость» и «мягкость» воды.
Соленость воды играет в жизни ряпушки большую роль. Плотность воды зависит от количества растворенных солей, следовательно условия плавания зависят от солености. Газовый режим также зависит от солености. Чем больше растворено в воде солей, тем меньше в ней кислорода.
От состава и количества растворенных в воде минеральных солей зависит развитие одноклеточных водорослей – пищи для беспозвоночных животных, а последние служат пищей для рыб.
Солевой состав воды оказывает на жизнь европейской ряпушки прямое влияние. Так, например, фосфор и кальций, имеющие важное значение при формировании костной ткани и синтезе белков, ряпушка может получать не только из пищи, но и непосредственно из воды. Магний, калий, натрий, серу, железо, медь, йод, фтор, молибден и другие химические элементы, необходимые для нормального роста и развития, они могут также получать из воды. Растворенные в воде минеральные соли поддерживают у ряпушки постоянное осмотическое давление, обеспечивают работу всех внутренних органов: всасывание в кровь через стенки кишечника питательных веществ, а также выделение продуктов обмена.
Различные виды рыб неодинаково реагируют на соленость воды. В связи с этим одних рыб, которые переносят значительное колебание солености воды, принято называть эвригалинными, а других рыб, не выдерживающих сильных изменений в количестве растворенных в воде солей, – стеногалинными. Европейская ряпушка относится к эвригалинным.
Газовый состав воды. Водная поверхность, соприкасающаяся с атмосферным воздухом, поглощает и растворяет азот, кислород и диоксид углерода (углекислоту и углекислый газ). Эти газы проникают в нижние слои воды и играют важную роль в жизни водных организмов. На жизнь европейской ряпушки они оказывают как прямое, так и косвенное влияние.
Содержание в воде растворенного кислорода изменяется в зависимости от температуры (с ее понижением повышается растворимость кислорода, и наоборот), атмосферного давления (чем выше давление тем больше растворимость кислорода), интенсивности перемешивания водных слоев (на больших глубинах кислорода меньше, чем в поверхностных слоях), от солености (чем выше соленость, тем меньше растворяется кислорода). Количество растворенного в воде кислорода также обусловлено биохимическими процессами, происходящими в самом водоеме (интенсивности фотосинтеза, поглощения кислорода и выделения углекислого газа). Растворенный в воде кислород необходим рыбам для дыхания. Рыбы по-разному реагируют на количество растворенного в воде кислорода. Одни нуждаются в очень высоком его содержании, к ним относится европейская ряпушка, другие менее требовательны, а третьи удовлетворяются ничтожным количеством (Иванов, 1988).
Количество потребляемого рыбами кислорода зависит от возраста рыб, степени зрелости половых продуктов и интенсивности питания. Для каждого вида существует как бы определенный порог содержания кислорода в воде, ниже которого они становятся вялыми, плохо питаются и, наконец, погибают. Так для европейской ряпушки содержание кислорода в воде ниже 1 – 3,5 мл/л приводит к удушью (Вавилкин, 1985).
Активная реакция среды. Этот показатель зависит от растворенных в воде различных химических веществ и определяется концентрацией в ней водородных ионов. Колебания активной реакции среды (рН) в водоеме бывает суточные, сезонные и годовые. Большое воздействие на значение рН оказывают кислород и диоксид углерода. Дыхание животных организмов и процессы гниения, происходящие в водоеме, уменьшая количество растворенного в воде кислорода и увеличивая содержание диоксида углерода, способствует снижению активной реакции среды.
При массовом развитии в водоеме растительных организмов, потребляющих из воды в светлое время суток диокси