Характеристика общих свойств микроорганизмов
1. Характеристика схожести и отличия прокариотических и эукариотических клетокОбоснование того, что прокариотный и эукариотный типы клеточной организации являются наиболее существенной границей, разделяющей все клеточные формы жизни, связано с работами Р. Стейниера (R. Stanier, 1916—1982) и К. ван Ниля, относящимися к 60-м гг. Поясним разницу между прокариотами и эукариотами. Клетка — это кусочек цитоплазмы, отграниченный мембраной. Последняя под электронным микроскопом имеет характерную ультраструктуру: два электронно-плотных слоя каждый толщиной 2,5—3,0 нм, разделенных электронно-прозрачным промежутком. Такие мембраны получили название элементарных. Обязательными химическими компонентами каждой клетки являются два вида нуклеиновых кислот (ДНК и РНК), белки, липиды, углеводы. Цитоплазма и элементарная мембрана, окружающая ее, — непременные и обязательные структурные элементы клетки. Это то, что лежит в основе строения всех без исключения клеток. Изучение тонкой структуры выявило существенные различия в строении клеток прокариот (бактерий и цианобактерий) и эукариот (остальные макро- и микроорганизмы).
Прокариотная клетка отличается тем, что имеет одну внутреннюю полость, образуемую элементарной мембраной, называемой клеточной, или цитоплазматической (ЦПМ). У подавляющего большинства прокариот ЦПМ — единственная мембрана, обнаруживаемая в клетке. В эукариотных клетках в отличие от прокариотных есть вторичные полости. Ядерная мембрана, отграничивающая ДНК от остальной цитоплазмы, формирует вторичную полость. Наружные мембраны хлоропластов и митохондрий, окружающие заключенные в них функционально специализированные мембраны, играют аналогичную роль. Клеточные структуры, ограниченные элементарными мембранами и выполняющие в клетке определенные функции, получили название органелл. Ядро, митохондрий, хлоропласты — это клеточные органеллы. В эукариотных клетках помимо перечисленных выше есть и другие органеллы.
В клетках прокариот органеллы, типичные для эукариот, отсутствуют. Ядерная ДНК у них не отделена от цитоплазмы мембраной. В цитоплазме находятся функционально специализированные структуры, но они не изолированы от цитоплазмы с помощью мембран и, следовательно, не образуют замкнутых полостей. Эти структуры могут быть сформированы и мембранами, но последние не замкнуты и, как правило, обнаруживают тесную связь с ЦПМ, являясь результатом ее локального внутриклеточного разрастания. В клетках прокариот есть также образования, окруженные особой мембраной, имеющей иное по сравнению с элементарной строение и химический состав.
Таким образом, основное различие между двумя типами клеток — существование в эукариотной клетке вторичных полостей, сформированных с участием элементарных мембран. Сопоставление некоторых черт клеточной организации прокариотных и эукариотных организмов представлено в таблица 1. Сопоставление некоторых черт прокариотной и эукариотной клеточной организации:
| Признак | Прокариотная клетка | Эукариотная клетка |
| Организация генетического материала | нуклеоид (ДНК не отделена от цитоплазмы мембраной), состоящий из одной хромосомы; митоз отсутствует | ядро (ДНК отделена от цитоплазмы ядерной оболочкой), содержащее больше одной хромосомы; деление ядра путем митоза |
| Локализация ДНК | в нуклеоиде и плазмидах, не ограниченных элементарной мембраной | в ядре и некоторых органеллах |
| Цитоплазматические органеллы | отсутствуют | имеются |
| Рибосомы в цитоплазме | 70S-типа | 80S-типа |
| Цитоплазматические органеллы | отсутствуют | имеются |
| Движение цитоплазмы | отсутствует | часто обнаруживается |
| Клеточная стенка (там, где она имеется) | в большинстве случаев содержит пептидогликан | пептидогликан отсутствует |
| Жгутики | нить жгутика построена из белковых субъединиц, образующих спираль | каждый жгутик содержит набор микротрубочек, собранны в группы: 2·9-2 |
В связи с тем что прокариотная и эукариотная организация клеток принципиально различна, было предложено только на основании этого признака выделить все прокариоты в особое царство. Р. Меррей (R. Murray) в 1968 г. предложил все клеточные организмы разделить на две группы по ТИПУ их клеточной организации: царство Prokaryotae, куда вошли все организмы с прокариотным строением клетки, н царство Eukaryotae, куда включены все высшие протисты, растения и животные.
Р. Виттэкер (R. Whittaker) предложил схему, по которой все живые организмы, имеющие клеточное строение, представлены разделенными на пять царств (рис. 2). Такая система классификации живого мира отражает три основных уровня его клеточной организации: Monera включает прокариотные организмы, находящиеся на самом примитивном уровне клеточной организации; Protista — микроскопические, в большинстве своем одноклеточные, недифференцированные формы жизни, сформировавшиеся в результате качественного скачка в процессе эволюции, приведшего к возникновению эукариотных клеток; многоклеточные эукариоты представлены в свою очередь тремя царствами Plantae, Fungi и Animalia.
Три последние таксономические группы различаются по способу питания: фототрофный тип питания за счет процесса фотосинтеза характерен для растений (Plantae): грибы (Fungi) в основном характеризуются осмотрофным типом питания, т. е. питанием растворенными органическими веществами; животные (Animalia) осуществляют голозойное питание, заключающееся в захватывании и переваривании твердой пищи. Способы питания, специфические для растений и грибов, возникли в процессе эволюции на уровне Monera. На уровне Protista они получили свое дальнейшее развитие; здесь же сформировался третий тип питания — голозойный.
Не берясь судить о целесообразности деления живой природы на пять или шесть царств, можно с определенностью утверждать, что обособление прокариотных микроорганизмов в отдельное царство Prokaryotae правомерно, поскольку основано на принципиальных различиях в структуре прокариотных и эукариотных клеток, т. е. тех единиц, из которых построены все клеточные формы жизни.
2. Характеристика морфологических и структурных признаков бактерий Firmicutes и gracilicutes
Firmicutes - тип бактерий, большинство из которых грамм-положительные. Некоторые (класс Mollicutes), совсем не имеют клеточной стенки и не красятся за Граммом, но не имеют и внешней мембраны, найденной в других грамм-отрицательных формах. Сначала к Firmicutes включали всех грамм-положительных бактерий, но сейчас они ограничиваются основной группой связанных форм, так называемой группой низкого содержимого G+C, в отличие от Actinobacteria. Они имеют круглые клетки, которые называют коками (хотя некоторые представители имеют палочковидные и спиральные формы).
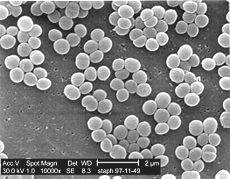
Електронна мікрофотографія колонії
Staphylococcus aureus
Много Firmicutes образовывают эндоспоры, которые есть очень стойкими к высушиванию и могут выдерживать экстремальные условия. Они найдены в разных окружающих средах, некоторые из них - известные патогены. Члены одной семьи, Heliobacteriaceae, получают энергию через фотосинтез.
Группа традиционно делилась на Clostridia (преимущественно анаэробы), Bacilli (облигатные или факультативные аэробы), и Mollicutes. На молекулярных деревьях первые две группы часто показываются как парафилетические или полифилетические к их основным родам, Clostridium и Bacillus, хотя поддержка этих данных слабая. Возможно, позднее эти группы будут пересмотрены.
Наиболее известные роды Firmicutes включают:
Bacilli, ряд Bacillales
Bacillus
Listeria
Staphylococcus
Bacilli, ряд Lactobacillales
Enterococcus
Lactobacillus
Lactococcus
Leuconostoc
Streptococcus
Clostridia
Acetobacterium
Clostridium
Eubacterium
Heliobacterium
Heliospirillum
Sporomusa
Mollicutes
Mycoplasma
Spiroplasma
Ureaplasma
Erysipelothrix
Способность образовывать более или менее терморезистентные споры, за немногими исключениями, ограничивается группой палочек, большинство из которых имеет перитрихально расположенные жгутики. Это грам-положительные бактерии. Аэробные и факультативно-анаэробные палочки сведены в роды Bacillus, Sporolactobacillus и Sporosarcina, а анаэробные роды Clostridium и Desulfotomaculum. Многие спорообразую-шие виды хорошо известны благодаря своим биохимическим особенностям. Здесь будут рассмотрены лишь некоторые представители больших родов Bacillus и Clostridium.
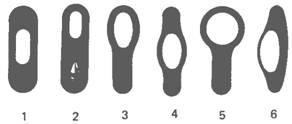
Рис. 2. Схематическое изображение типичных форм спорообразующих клеток. 1 -центрально расположенная спора, которая не увеличивает размера материнской клетки (Bacillus megaterium); 2-терминальная спора, не увеличивающая размеров материнской клетки (Bacillus thuringiensis), с белковыми включениями; 3 - терминальная спора, расширившая материнскую клетку булавовидно (Bacillus macerans); 4-центральная спора, придающая материнской клетке веретенообразную (клостридиеподобную) форму (Bacillus polymyxa); 5-круглая терминальная спора, придавшая материнской клетке форму барабанной палочки (плектри-диальную форму) (Bacillus sphaericus); 6-латеральная спора; материнская клетка увеличилась в размерах, приняв форму веретена (Bacillus laterosporus).
Аэробные спорообразующие бактерии. Бактерии этого типа обитают в почве. Многие бациллы образуют цепочки клеток или нити. По форме спор и материнских клеток (рис. 3.6) бациллы могут быть разделены на три группы:
(I) Споры овальные или цилиндрические, не толще материнской клетки. Таковы споры большинства бацилл (В. megaterium, В. cereus, В. subtilis, В. licheniformis, B. anthracis, B. thuringiensis).
(II) Овальные споры шире материнской клетки; они «раздувают» клетку изнутри в ходе споруляции (В. polymyxa, В. macerans, В. stearothermophilus, В. circulans).
(III) Почти круглая спора в набухшем конце материнской клетки (В. pasteurii).
I. Bacillus megaterium с клетками величиной 2x5 мкм является гигантом среди эубактерий. Несколько меньше В. cereus; к этому виду теперь причисляют также вариант, получивший название «mycoides» за грибовидный рост на поверхности агара (В. cereus var. mycoides). Существуют «право- и левозакрученные» штаммы, но общий вид колоний очень типичен. В близком родстве с В. cereus находятся возбудитель сибирской язвы В. anthracis, который не имеет жгутиков и окружен капсулой из глутаминовой кислоты, и патогенная для насекомых бактерия В. thuringiensis. Вид Bacillus subtilis, получивший название «сенной палочки» (так как накопительные культуры получают из сенного экстракта), а также В. licheniformis образуют полипептидные антибиотики. Вторая из названных бацилл способна расти, получая энергию не только за счет кислородного дыхания, но также за счет брожения и анаэробного нитратного дыхания.
П. Бактерия В. polymyxa (ранее называвшаяся В. asterosporus) получила нынешнее свое название из-за того, что образует большое количество слизи (а также боченкообразные, в поперечном сечении звездчатые споры). Как и В. licheniformis, она образует 2,3-бутандиол. В. stеarothermophilus - выраженный термофил (температурный оптимум для роста 50-65°С).
III. Вид В. pasteurii известен как классический пример бациллы, разлагающей мочевину; он конститутивно синтезирует уреазу, гидролизует мочевину до С02 и аммиака и приспособлен к высоким значениям рН. По своей физиологии с ним сходна Sporosarcina ureae, которая в морфологическом отношении, собственно, относится к сарцинам, но по основным физиологическим признакам должна быть отнесена к бациллам (аэроб; образует терморезистентные споры, содержащие дипиколино-вую кислоту).
Анаэробные спорообразующие бактерии. Для роста этих бактерий кислород не нужен. Виды, объединенные в род Clostridium, как правило, не имеют цитохромов и каталазы. Из-за высокого содержания флавиновых ферментов многие клостридии при контакте с кислородом воздуха образуют перекись водорода, которая оказывает на их клетки токсическое действие. Ввиду того что сульфатредуцирующие бактерии, образующие споры, содержат протогеминовые пигменты, их выделили из рода Clostridium в новый род Desulfotomaculum (D. nitrificans, D. orientis, D. ruminis). У анаэробных спорообразующих бактерий спора обычно значительно шире, чем вегетативная клетка. Материнская клетка в зависимости от положения споры может иметь различную форму.
Клостридии сбраживают большое число субстратов, включая полисахариды, белки, аминокислоты и пурины. По предпочитаемым субстратам можно различать сахаролитические клостридии (например, Clostridium butyricum, С. acetobutylicum, С. cellulosae-dissolvens), пептолитические клостридии (С. histolyticum, С. sporogenes, С. tetani, С. botulinum) и клостридии, разлагающие мочевую кислоту (С. acidi-urici). Продуктами брожения являются бутират, бутанол, ацетон, 2-пропанол и во многих случаях большие количества газа (Н2, С02). С. Pasteurianum и многие другие сахаролитические клостридии фиксируют молекулярный азот. С. aceticum превращает фруктозу или смесь С02 и молекулярного водорода в ацетат.
В качестве дополнения следует указать на Oscillospira guilliermondii--необычно крупную (5 х 100 мкм) анаэробную бактерию, разделяющуюся на клетки и образующую споры. Ее часто находят в слепой кишке морской свинки и в рубце жвачных.
Протеобактерии (proteobacteria) это одна из основных групп бактерий. Они насчитывают большое количество патогенов, таких как кишечная палочка (E.coli), сальмонела (Salmonella), вибрион (Vibrio), чумная палочка (Yersinia pestis), Helicobacter pylori и много других видов. Другие есть свободно живучими организмами, и насчитывают много бактерий, отвечающих за фиксацию азота. Как и для других бактерий, группа определяется по последовательностью рибосомной РНК (16S ррнк), и названная за именем греческого бога Протея (также как и один из родов бактерий сред протеобактерий), который мог изменять свою форму, в результате большого разнообразия форм, найденных в этой группе.
Все протеобактерии грамм-отрицательные, с внешней мембраной преимущественно построенной из полисахаридов. Многие из них двигаются применяя джгутики, но некоторые недвижимые, а другие викорустовують ворсинки или другие механизмы бактериального скольжения. Последние включают миксобактерий, уникальную группу бактерий, которые могут агрегуватися чтобы создать многоклеточные плодовые тела. Между протеобактерий также существует большое разнообразие типов метаболизма. Большинство членов являются факультативными или облигатними анаеробами и гетерофами, но также существуют множество исключения. Разные виды, не будучи родственными, превращают энергию света, применяя фотосинтез. Также среди протеобактерий существуют пурпурные бактерии, названные так за их красновату пигментацию.
Альфа-протеобактерии охватывают большинство фототрофных видов, но также несколько видов, метаболизующих C 1-вещества, симбиоты с растениями (ризобии) и животными, и группу опасных патогенов, Rickettsiaceae. Больше того, ученые считают, что предтечи митохондрий клеток эукариотов также возникли из этой группы.
Бета-протеобактерии содержат несколько групп аэробных или факультативных бактерий, которые часто очень разнообразные за своими возможностями метаболизма, крот того, хемолоитотрофные роды (т.е., окисляющий аммиак род Nitrosomonas) и фототрофов (роды Rhodocyclus и Rubrivivax). Многие из них найденные только в естественных образцах, например, видбросах и грунту. Этот класс содержит и некоторое количество патогенных видов: Neisseriaceae (gonorrhoe и meningoencephalitis) и виды рода Burkholderia.
Гамма-протеобактерии охватывают несколько клинично и научно важных групп of бактерий, таких как Enterobacteriaceae, Vibrionaceae и Pseudomonadaceae. Значительное число важных патогенив принадлежит к этому классу, т.е. Salmonella (вызывает энтерит и тиф), Yersinia (чума), Vibrio (холера), Pseudomonas aeruginosa (инфекции легких госпителизованых или клиничных пациентов с кистозным фиброзом).
Дельта-протеобактерии охватывают подраздел большей частью аэробных бактерий, бактерий образующих плодовые тела миксобактерии, и подраздел строго аэробных видов, который содержит большинство известных сульфат- (Desulfovibrio, Desulfobacter, Desulfococcus, Desulfonema, и т.п.) и серо-восстанавливающие бактерии (e.g. Desulfuromonas) и также несколько других анаеробных бактерии с другой физиологией (т.е. зализо-видновлюючи Geobacter и синтропичные виды: Pelobacter и Syntrophus).
Эпсилон-протеобактерии содержат лишь несколько видов, преимущественно вигнети и спиральные Wolinella, Helicobacter, и Campylobacter. Они все насиляють травный тракт животных и человека и служат как симбиоти (Wolinella в коровах) или патогени (Helicobacter в желудке, Campylobacter в двнадцатиперсний кишке).
Планктомицети - облигатно аэробные водные бактерии, найденные в образцах соленой и пресной воды. Они размножаются почкованием. По структуре, организмы этой группы яйцевидные и имеют структуру непепродуктивного конца, которая помогает им соединяться на протяжении почкования, так называемое стебель.
Организмы, которые принадлежат к этой группе не имеют пептидогликанив в клитиннии стенке. Пептидогликан - важный гетерополимер, присутствующий клеточных стенках большинства бактерий, который служат защитным компонентом. Вместе с тем стенки планктомицетов составлены с гликопротеинив богатых на глутамин. Планктомицети имеют внутренние структуры, более сложные, чем обычно ожидаются в прокариотах. Тогда как они не имеют клидинного ядра такой структуры как в эукариот, ядерный материал иногда может быть окружен в двойной мембране. В дополнение к этому нуклеоиду, эти бактерии содержат два других отдела, отделенных мембраной, пирреллюлозому или рибоплазму, которая содержит рибосомы и связанные белки, и свободный от рибосом парафоплазму (Glockner, 2003).
3. Характеристика общих свойств микроорганизмов
Микроорганизмы - это организмы, невидимые невооруженным глазом из-за их незначительных размеров. Этот критерий - единственный, который их объединяет. В остальном мир микроорганизмов еще более разнообразен, чем мир макроорганизмов. Согласно современной систематике, микроорганизмы относятся к трем царствам:
Vira - к ним относятся вирусы;
Eucariotae - к ним относятся простейшие и грибы;
Procariotae - к ним относятся истинные бактерии, риккетсии, хламидии, микоплазмы, спирохеты, актиномицеты.
Количество и жизнедеятельность микробов зависят от условий существования (питания, температуры, влажности).
По характеру питания микробы делятся на автотрофные, питающиеся минеральными веществами, и гетеротрофные, питающиеся готовыми органическими соединениями. Гетеротрофные микробы делятся на сапрофиты (метатрофы), разлагающие органические вещества в природе и вызывающие порчу пищевых продуктов, и паразиты (паратрофы), развивающиеся в теле других организмов и питающиеся сложными органическими веществами. К группе паразитов относятся разнообразные возбудители заболеваний человека и животных.
По типу дыхания микробы делятся на аэробы, развивающиеся только при доступе кислорода воздуха, и анаэробы, не нуждающиеся в кислороде воздуха. Анаэробные микробы подразделяются на облигатные, для которых кислород вреден, и факультативные, которые могут жить как при доступе воздуха, так и без него Температура является одним из наиболее важных факторов, влияющих на жизнедеятельность микробов.
По отношению к температуре микробы подразделяют на три группы мезофилы, психрофилы и термофилы.
Большинство микробов, находящихся в стадии активного размножения (вегетативная стадия), погибает при температуре около 70° С за 1—5 мин. Споры некоторых бактерий выдерживают кипячение в течение нескольких часов. Во влажной среде споры бактерий погибают при 120° С через 20—30 мин, а в с^хой — при 160—170° С через 1—2 ч Споры большинства дрожжей и плесеней менее устойчивы к воздействию высоких температур, чем споры бактерий и быстро погибают при нагревании до 65—80° С
Влияние температуры на жизнедеятельность микробов обусловливает возможность хранения пищевых продуктов (рыба, мясо и др.) при пониженных температурах, замедляющих размножение микробов и угнетающих деятельность ферментов. Наблюдения показывают, что количество микробов, погибших при замораживании продуктов, нередко достигает 80—90% от их первоначального содержания. Микроорганизмы, оставшиеся в живых, вначале инактивируются холодом, но при дальнейшем хранении охлажденного или замороженного продукта при температуре не ниже минус 8° С их жизнедеятельность постепенно восстанавливается Губительно действуют на микробы повторное замораживание и оттаивание продукта.
Большое влияние на жизнедеятельность микробов оказывает влажность среды. В результате высушивания продукта останавливается развитие многих видов содержащихся в нем микробов, так как при отсутствии воды они не могут питаться. Так, минимум содержания влаги в среде обитания для развития бактерии составляет 30%, а для многих плесеней — около -13% . Споры некоторых плесневых грибов сохраняют способность к прорастанию при отсутствии влаги в течение нескольких лет.
4. Особенности строения муреина у бактерий.
Опорный скелет бактериальной стенки тоже состоит в значительной степени из однородного полимера-пептидогликана муреина. Эта макромолекула - гетерополимер, построенный из цепочек, в которых чередуются остатки N-ацетилглюкозамина и N-ацетилмурамовой кислоты (N-ацетилглюкозаминлактата), соединенные между собой рЧ,4-глико-зидными связями.
Такие неразветвленные гетерополимерные цепи образуют основу муреина. Остатки мурамовой кислоты через лактильные группы соеди нены пептидной связью с аминокислотами. К типичным аминокислотам муреина относятся L-аланин, D-глутаминовая кислота, .мезодиаминопи-мелиновая кислота или L-лизин и D-аланин. Диаминокислоты мезо-(или LL-) диаминопимелиновая кислота и L-лизин играют большую роль в межмолекулярных сшивках, так как образуют пептидные связи с участием обеих аминогрупп и, таким образом, могут связать две гетерополимерные цепи между собой. Место диаминопимелино-вой кислоты или лизина могут занять орнитин или диаминомасляная кислота. Пептидными мостиками гетерополимерные цепи связаны между собой в мешкообразную гигантскую молекулу -муреиновый мешок.
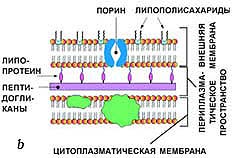
Муреиновый мешок выполняет функцию опорного каркаса клеточной стенки. На нем откладываются и его инкрустируют различные другие вещества. По строению этого каркаса, а также по содержанию других веществ в клеточной стенке грам-положительные бактерии отличаются от грам-отрицательных.
Существуют 2 типа строения клеточной стенки:
· у бактерий первого типа пептидогликан муреин составляет до 90% массы клеточной стенки и образует многослойный (до 10 слоев) каркас, при этом он ковалентно связан с тейхоевыми кислотами. Такие бактерии при окраске по методу Грама прочно удерживают комплекс генцианового фиолетового и йода; они окрашиваются в сине-фиолетовый цвет и называютсяграмположителъными:
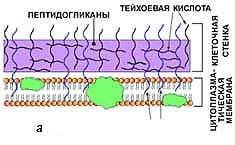
· у бактерий со вторым типом строения клеточной стенки поверх 2—3 слоев пептидогликана муреина располагается слой липополисахаридов. Эти бактерии при окраске по методу Грама неспособны прочно удерживать комплекс генцианового фиолетового и йода и соответственно обесцвечиваются спиртом, прокрашиваясь дополнительным красителем — фуксином — в розово-красный цвет. Они называются грамотрицательными.
5. Характеристика микроорганизмов по способам питания
Объединить типы конструктивного и энергетического метаболизма можно в следующей таблице:
| Способы существования живых организмов (матрица Львова) | ||||
| Источник энергии | Донор электрона | Источник углерода | Название способа существования | Представители |
| ОВР | Неорганические соединения | Углекислый газ | Хемолитоавтотрофия | Нитрифицирующие, тионовые, ацидофильные железобактерии |
| Органические соединения | Хемолитогетеротрофия | Метанообразующие архебактерии, водородные бактерии | ||
| Органические вещества | Углекислый газ | Хемоорганоавтотрофия | Факультативные метилотрофы, окисляющие муравьиную кислоту бактерии | |
| Органические соединения | Хемоорганогетеротрофия | Большинство прокариот, из эукариот: животные, грибы, человек | ||
| Свет | Неорганические соединения | Углекислый газ | Фотолитоавтотрофия | Цианобактерии, пурпурные, зелёные бактерии, из эукариот: растения |
| Органические соединения | Фотолитогетеротрофия | Некоторые цианобактерии, пурпурные, зелёные бактерии | ||
| Органические вещества | Углекислый газ | Фотоорганоавтотрофия | Некоторые пурпурные бактерии | |
| Органические вещества | Фотоорганогетеротрофия | Галобактерии, некоторые цианобактерии,пурпурные, зелёные бактерии | ||
Из таблицы видно, что разнообразие типов питания прокариот в разы больше чем у эукариот (последние способны лишь к хемоорганогетеротрофии и фотолитоавтотрофии).
6. Химическое строение и структурная организация вирусов, морфология, особенности взаимодействия с клеткой-хозяином.
Вирусы не имеют клеточного строения. Каждая вирусная частица состоит из расположенного в центре носителя генетической информации и оболочки. Генетический материал представляет собой короткую молекулу нуклеиновой кислоты, это образует сердцевину вируса. Нуклеиновая кислота у разных вирусов может быть представлена ДНК или РНК, причем эти молекулы могут иметь необычное строение: встречается однонитчатая ДНК и двух нитчатая РНК.
Оболочка называется капсид. Она образована субъединицами – капсомерами, каждый из которых состоит из одной или двух белковых молекул. Число капсомеров для каждого вируса постоянно (в капсиде вируса полиомиелита их 60, а у вируса табачной мозаики – 2130). Иногда нуклеиновая кислота вместе с капсидом называется нуклеокапсидом. Если вирусная частица кроме капсида, больше не имеет оболочки, её называют простым вирусом, если имеется ещё одна – наружная, вирус называется сложным. Наружную оболочку также называют суперкапсидом, генетически она не принадлежит вирусу, а происходит из плазматической мембраны клетки-хозяина и формируется при выходе собранной вирусной частицы из инфицированной клетки.
У каждого вируса капсомеры капсида располагаются в строго определённом порядке, благодаря чему возникает определённый тип симметрии. При спиральной симметрии капсид приобретает трубчатую (вирус табачной мозаики) или сферическую (РНК-содержащие вирусы животных) форму. При кубической симметрии капсид имеет форму икосаэдра (двадцатигранника), такой симметрией обладают изометрические вирусы. В случае комбинированной симметрии капсид обладает кубической формой, а расположенная внутри нуклеиновая кислота уложена спирально. Правильная геометрия капсида даже позволяет вирусным частицам совместно образовывать кристаллические структуры.
Непременным компонентом вирусной частицы является одна из двух нуклеиновых кислот, белок и зольные элементы. Эти три компонента являются общими для вирусов, тогда как остальные двалипоиды и углеводы - входят в состав далеко не всех вирусов.
Вирусы, состоящие только из белка нуклеиновой кислоты и зольных элементов, чаще всего принадлежат к группе простых вирусов, лишенных дифференциации, собственных ферментов или каких-либо специализированных структур - вирусы растений, некоторые вирусы животных и насекомых. В то же время практически все бактериофаги, которые по химическому составу, принадлежат к группе минимальных вирусов, на самом деле являются очень сложными и высокодифференцированными структурами.
Вирусы, в состав которых наряду с белком и нуклеиновой кислотой входят также липоиды и углеводы, как правило, принадлежат к группе сложно устроенных вирусов. Большая часть вирусов этой группы паразитирует на животных.
Белок всех исследованных до настоящего времени вирусов построен из обычных аминокислот, принадлежащих к естественному L-ряду. Соотношение аминокислот в вирусных белках достаточно близко к таковому в белках животных, бактерий и растений.
Вирусные белки не содержат обычно большого количества основных аминокислот (аргинина, муцина). Не учитывая нейтральных аминокислот, можно сказать, что в вирусном белке преобладают кислые дикарбоновые кислоты. Это справедливо для вирусов с низким и высоким содержанием нуклеиновых кислот.
Молекулы вирусных ДНК могут быть линейными или кольцевыми, двух цепочечными или одно цепочечными по всей своей длине или же одно цепочечными только на концах. Кроме того, выяснилось, что большинство нуклеотидных последовательностей в вирусном геноме встречается лишь по одному разу, однако на концах могут находиться повторяющиеся, или избыточные участки. Помимо различий в форме молекулы и в структуре концевых участков вирусных ДНК существуют также различия в величине генома.
Исследования вирусной РНК составили один из самых значительных вкладов вирусологии в молекулярную биологию. Тот факт, что у вирусов растений реплицируемая генетическая система состоит только из РНК, ясно показал, что и РНК способна сохранять генетическую информацию. Была установлена инфекционность РНК вируса табачной мозаики, и выяснилось, что для инфекции необходима вся ее молекула.
Размеры вирионов РНК - вирусов сильно варьируют - от 7.106 до 2.108 дальтон, однако размеры РНК и, следовательно, объем содержащейся в ней информации различаются в значительно меньшей степени.
Четверым компонентом, обнаруживаемым иногда в очищенных вирусных препаратах, являются углеводы (в количестве, превышающем содержание сахара в нуклеиновой кислоте). Глюкоза и гентибиоза обнаружена в составе некоторых фагов. Помимо этих углеводов, в составе бактериофагов могут быть и другие полисахариды. Единственная группа вирусов, в которой наличие углеводов точно доказано - вирусы животных. В составе элементарных телец вируса гриппа и классической чумы птиц находятся до 17 % углеводов.
Наиболее важный из таких компонентов двойной слой липидов, образующий основную массу наружной оболочки у тех вирусов, у которых она имеется. Полагают, что липиды оболочек просто заимствуются из плазматической мембраны клетки-хозяина и поэтому, строго говоря, не могут считаться вирус специфическими.
Высокоочищенные препараты вирионов содержат ряд низкомолекулярных компонентов. У бактериофагов и вирусов животных и растений обнаружены полиамины. Возможно, что их физиологическая функция состоит в нейтрализации отрицательного заряда нуклеиновой кислоты. Например, вирус герпеса содержит достаточно спермина, чтобы нейтрализовать половинку вирусной ДНК, а в вирусной оболочке присутствует спермидин.
В состав некоторых вирусов растений (морщинистости турнепса, крапчатости фасоли, табачной мозаики) входит бис амин.
Вирусная частица - это инертная статическая форма вируса. Когда вирионы находятся вне клетки, они не размножаются и в них не происходит никаких метаболических процессов. Все динамические события начинаются лишь тогда, когда вирус проникает в клетку. Даже у многоклеточного хозяина решающие события при вирусной инфекции происходят на клеточном уровне. Распространение вируса совершается в результате повторных циклов взаимодействия вируса с клетками и рассеяния вирионов во внеклеточной среде.
В зараженных вирусом клетках происходит глубокая перестройка вирусного материала, а часто также и компонентов клетки-хозяина. Возникает новая система - комплекс вирус-клетка. Репродукция вирусов – процесс многоэтапный, который можно разделить на семь стадий:
1.Адсорбция. Это процесс прикрепления вирусов к поверхности восприимчивой клетки. Вначале вирионы адсорбируются посредством электростатического взаимодействия или за счёт ван-дер-ваальсовых сил. Эта стадия обратима: вирус можно отделить обычным встряхиванием.
2.Инъекция. Связана с введением (инъекцией) в клетку инфекционной нуклеиновой кислоты вируса (как у фагов) или проникновением в клетку целой вирусной частицы с последующим «раздеванием» вируса от белковой оболочки и высвобождением инфекционной нуклеиновой кислоты.
3.Депротеинизация. В ходе её происходит освобождение носителя генетической информации вируса – его нуклеиновой кислоты. У бактериофагов этот процесс совпадает с предыдущей стадией.(1)
4.Репликация вирусных молекул нуклеиновой кислоты. Репликация идёт за счет нуклеотидов, накопленных в клетке хозяина.
5.Синтез вирус специфических структурных белков и ферментов. Процесс синтеза идёт в рибосомах клетки хозяина.
6.Сборка (самоорганизации) вирусных частиц. Для этого необходимо, чтобы концентрация компонентов вириона достигла высокого (критического) уровня. Компоненты вирусной частицы синтезируются раздельно и в разных частях клетки.(2) Сначала происходит комплексирование нуклеиновых кислот с частью белков и образование нуклеопротеидов. Последние покрываются оболочками. В состав этих оболочек входят часто некоторые компоненты клеточной мембраны.
Лизис. У бактерий распад клеток происходит под влиянием ферментов фага, а у клеток высших организмов – путём выпячивания оболочки клеток и «выталкивания» вирусных частиц в окружающую среду.
Бактериофаги вирусы бактерий. Впервые описаны Ф. Туортом (1915), а термин был введён Ф. Д’Эреллем (1917).
Рассмотрим типичный бактериофаг на примере вируса кишечной палочки. Тело фага состоит из головки, от которой отходит полый стержень, окружённый чехлом из сократительного белка. Стержень заканчивается базальной пластинкой, на которой закреплены шесть нитей. Внутри головки находится ДНК.
Бактериофаг при помощи отростков прикрепляется к поверхности кишечной палочки и в месте соприкосновения с ней растворяет с помощью фермента клеточную стенку. После этого за счёт сокращения головки молекула ДНК фага впрыскивается через канал стержня в клетку. Примерно через 10-15 мин под действием этой ДНК перестраивается весь метаболизм бактериальной клетки, и она начинает синтезировать ДНК бактериофага, а не собственную. При этом синтезируется и фаговый белок. Завершается процесс появлением 200-1000 новых фаговых частиц, в результате чего клетка бактерии погибает.
Бактериофаги, образующие в заражённых клетках новое поколение фаговых частиц, что приводит к лизису (распаду) бактериальной клетки, называются вирулентными фагами.
Некоторые бактериофаги внутри клетки хозяина не реплицируются. Вместо этого их нуклеиновая кислота включается в ДНК хозяина, образуя с ней единую молекулу, способную к репликации. Такие фаги получили название умеренных фагов или профагов.
Прокариоты не дифференцируются в стволовые или специализированные клетки, а являются популяцией более или менее сходных клеток, которые продолжают размножаться, пока имеется соответствующая питательная среда. Поэтому взаимодействие фагов с бактериями происходит в бактериальной культуре циклически, пока не наступит некое равновесное состояние, которое определяется числом клеток-хозяев и вирусных элементов и скоростью их воспроизведения.
7. Понятие биологической системы, биологического объекта, гомеостаз.
Понятие открытой системы ввел Л. фон Берталанфи. Основные отличительные черты открытых систем - способность обмениваться со средой массой, энергией и информацией. К ним безусловно относятся биологические системы.
Одна из наиболее полных и интересных классификаций по уровням сложности предложена К. Боулдингом. Выделенные в ней уровни приведены в табл. 1.
| Тип системы | Уровень сложности |
Подобные работы:
Актуально:
|