Современная концепция гена
План.
1. Введение
2. Понятие «ген». Развитие представлений о нем
3. Структура генов. Классификация генов
4. Функции генов
5. Современные представления о генотипе.
6. Заключение
7. Список использованной литературы.
Введение.
Цель моего реферата – попытка раскрыть фундаментального понятия современной генетики – понятия «ген». Генетика – молодая наука, она начала развиваться только в XX веке. Генетика изучает законы двух фундаментальных свойств живых организмов - наследственности и изменчивости, лежащие в основе эволюции органического мира и деятельности человека по созданию новых сортов культурных растений, пород животных и штаммов микроорганизмов.
Наследственность – это свойство организма передавать свои признаки и особенности развития следующим поколениям.
Изменчивость – это свойство организмов приобретать новые признаки в процессе индивидуального развития.
Оба этих фундаментальных свойств организмов (и наследственность, и изменчивость) осуществляются генами. Гены хранят и передают информацию об организме последующим поколениям.
Понятие «ген». Развитие представлений о нем.
Ген – функциональная единица наследственного материала. Ген (от греч. genos — род, происхождение) – участок молекулы геномной нуклеиновой кислоты, характеризуемый специфической для него последовательностью нуклеотидов, представляющий единицу функции, отличной от функций других генов, и способный изменяться путем мутирования.
От гипотетических дискретных наследственных факторов до локализованных в хромосомах и молекулах ДНК генов. Долгое время ген рассматривали как минимальную часть наследственного материала (генома), обеспечивающую развитие определенного признака у организмов данного вида. Однако каким образом функционирует ген, оставалось неясным. Термин ген предложен В. Иогансеном в 1909 году, однако проникновение в его сущность связано с именем Г. Менделя, который еще в 1860-х гг. ввел термин «наследственный фактор» и на основе точных экспериментов сделал гениальные обобщения относительно свойств и поведения наследственных факторов при передаче информации от родителей потомкам, которые в последующем легли в основу теории гена. Это следующие фундаментальные свойства наследственных факторов – генов:
1) наличие альтернативных наследственных факторов для развития каждого конкретного признака организма (в современном представлении доминантный и рецессивный аллели гена).
2) Парность наследственных факторов, определяющих развитие признака (у диплоидного организма). Существенный вывод: наследуются не признаки, а от родителей к потомкам передаются вместе с гаметами гены. Из этих двух положений был развит принцип аллелизма.
3) Относительное постоянство гена.
Мендель не имел никаких сведений о местонахождении наследственных факторов в клетке, и тем более об их химической природе и механизме влияния на признак, т. е. наследственный фактор в начале 20 века выступал как условная единица наследственности.
Дальнейшая конкретизация представлений о гене связана с работами школы американского биолога Т. Х. Моргана. Введя в генетические исследования плодовую мушку-дрозофилу, удалось существенно увеличить разрешающую способность генетического анализа и на основе синтеза генетических и цитологических представлений доказать существование материальной структуры наследственности – хромосом, в которых локализованы гены.
Доказательствами хромосомной локализации генов явились: открытие генов, наследующихся сцеплено с полом (локализация генов в половых хромосомах, X или Y); сцепленное наследование группы признаков. Было показано наличие определенного числа групп сцепления генов, соответственно гаплоидному числу хромосом конкретного биологического вида. Кроме того, были получены генетические и цитологические доказательства кроссинговера – обмена генами между гомологичными хромосомами, приводящего к рекомбинации генов. Величина генетической рекомбинации (процент кроссинговера-перекреста) отражает расстояние между генами одной группы сцепления: чем дальше отстоят друг от друга гены, тем больше процент кроссинговера.
Таким образом, было доказано, что гены в хромосоме располагаются в линейном порядке, и каждый ген имеет свое определенное местоположение – локус. Соответственно открылась возможность построения плана взаимного расположения в хромосоме известных генов с указанием относительных расстояний между ними, выраженных в процентах перекреста (генетические карты) и идентифицировать местоположение гена в хромосоме (цитологические карты).
В 1945 г. Дж. Бидлом и Э. Татумом была сформулирована гипотеза, которую можно выразить формулой «Один ген - один фермент». Согласно этой гипотезе, каждая стадия метаболического процесса, приводящая к образованию в организме (клетке) какого-то продукта, катализируется белком-ферментом, за синтез которого отвечает один ген.
Позднее было показано, что многие белки имеют четвертичную структуру, в образовании которой принимают участие разные пептидные цепи. Поэтому формула, отражающая связь между геном и признаком, была несколько преобразована: «Один ген - один полипептид».
Изучение химической организации Э. Чаргаффом наследственного материала и процесса реализации генетической информации привело к формированию представления о гене как о фрагменте молекулы ДНК, транскрибирующемся в виде молекулы РНК, которая кодирует аминокислотную последовательность пептида или имеет самостоятельное значение (тРНК и рРНК).
Также ценные сведения о структуре ДНК дали результаты рентгеноструктурного анализа. Рентгеновские лучи, проходя через кристалл ДНК, претерпевают дифракцию, т.е. отклоняются в определенных направлениях. Степень и характер отклонения зависят от структуры самой молекулы. Анализ дифракционных рентгенограмм привел к заключению, что азотистые основания уложены на подобие стопки тарелок. Рентгенограммы позволили выявить в ДНК 3 главных периода: 0,34, 2 и 3,4, которые оказались размерами в модели ДНК, предложенной Дж.Уотсоном и Ф.Криком. 0,34 нм – расстояние между последовательными нуклеотидами, 2 нм – толщина цепи, 3,4 нм – расстояние между последовательными витками спирали.
В конце двадцатых годов советские генетики А. С. Серебровский и Н. П. Дубинин экспериментально показали, что ген не является единицей мутации, что он имеет сложную структуру: состоит из нескольких субъединиц, способных самостоятельно мутировать (ступенчатый аллелизм, или центровая теория гена). Весь ген (базиген) может состоять из отдельных центров, трансгенов, каждый из которых несет сходную функцию. Мутация может нарушать деятельность одного из трансгенов, не затрагивая других.
Несколько позже идея о сложном строении гена была подкреплена экспериментами по внутригенному кроссинговеру на дрозофиле по локусам lozenge, white и др. (работы Э. Льюиса, М. Грина и др.).
Таким образом, к 1950 году ген представлялся как участок хромосомы, контролирующий развитие определенного признака, имеющий определенную линейную протяженность и способный мутировать в разных участках и быть разделенным кроссинговером. Ген комплексен, так как его отдельные участки могут различаться по функциям, и в их совместной деятельности существует определенная субординация.
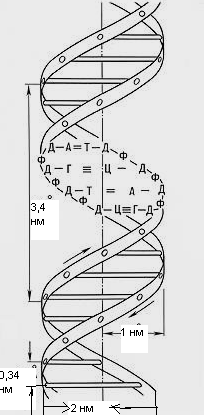 Схема участка ДНК.
Схема участка ДНК.
Структура генов.
Ген представляет собой последовательность нуклеотидов ДНК размером от нескольких сотен до миллиона пар нуклеотидов, в которых закодирована генетическая информация о первичной структуре белка (число и последовательность аминокислот). Для регулярного правильного считывания информации в гене должны присутствовать: кодон инициации, множество смысловых кодонов и кодон терминации. Три подряд расположенных нуклеотида представляют собой кодон, который и определяет, какая аминокислота будет располагаться в данной позиции в белке. Например, в молекуле ДНК последовательность оснований ТАС является кодоном для аминокислоты метионина, а последовательность ТТТ кодирует фенилаланин. В молекуле иРНК вместо тимина (Т) присутствует основание урацил (У). Таблица генетического кода во всех руководствах представлена именно символами иРНК. Из 64 возможных кодонов смысловыми являются 61, а три триплета — УАА, УАГ, УГА — не кодируют аминокислоты и поэтому были названы бессмысленными, однако на самом деле они представляют собой знаки терминации трансляции.
Для прокариот характерна относительно простая структура генов. Так, структурный ген бактерии, фага или вируса, как правило, контролирует одну ферментативную реакцию. Специфичным для прокариот является оперонная система организации нескольких генов. Гены одного оперона (участка генетического материала, состоящего из 1, 2 и более сцепленных структурных генов, которые кодируют белки (ферменты), осуществляющие последовательные этапы биосинтеза какого-либо метаболита; в оперон эукариот входит, как правило, 1 структурный ген; оперон содержит регуляторные элементы) расположены в кольцевой хромосоме бактерии рядом и контролируют ферменты, осуществляющие последовательные или близкие реакции синтеза (лактозный, гистидиновый и др. опероны).
Структура генов у бактеориофагов и вирусов в основном схожа с бактериями, но более усложнена и сопряжена с геномом хозяев. Например, у фагов и вирусов обнаружено перекрывание генов, а полная зависимость вирусов эукариот от метаболизма клетки-хозяина привела к появлению экзон-интронной структуры генов.
Эукариотические гены, в отличие от бактериальных, имеют прерывистое мозаичное строение. Кодирующие последовательности (экзоны) перемежаются с некодирующими (интронами). Экзон (от англ. ex(pressi)on — выражение, выразительность) - участок гена, несущий информацию о первичной структуре белка. В гене экзоны разделены некодирующими участками — интронами. Интрон (от лат. inter — между) - участок гена, не несущий информацию о первичной структуре белка и расположенный между кодирующими участками — экзонами. В результате структурные гены эукариот имеют более длинную нуклеотидную последовательность, чем соответствующая зрелая иРНК, последовательность нуклеотидов в которой соответствует экзонам. В процессе транскрипции информация о гене списывается с ДНК на промежуточную иРНК, состоящую из экзонов и интронов. Затем специфические ферменты — рестриктазы — разрезают эту про-иРНК по границам экзон-интрон, после чего экзонные участки ферментативно соединяются вместе, образуя зрелую иРНК (так называемый сплайсинг). Количество интронов может варьировать в разных генах от нуля до многих десятков, а длина — от нескольких пар оснований до нескольких тысяч.
Ген может кодировать различные РНК-продукты путем изменения инициирующих и терминирующих кодонов, а также альтернативного сплайсинга. Альтернативная экспрессия гена осуществляется и путем использования различных сочетаний экзонов в зрелой иРНК, причем полипептиды, синтезированные на таких иРНК, будут различаться как по количеству аминокислотных остатков, так и по их составу.
Наряду со структурными и регуляторными генами обнаружены участки повторяющихся нуклеотидных последовательностей, функции которых изучены недостаточно, а также мигрирующие элементы (мобильные гены), способные перемещаться по геному. Найдены также так называемые псевдогены у эукариот, которые представляют собой копии известных генов, расположенные в других частях генома и лишенные интронов или инактивированные мутациями.
Классификация генов.
Накопленные знания о структуре, функциях, характере взаимодействия, экспрессии, мутабильности и других свойствах генов породили несколько вариантов классификации генов.
По месту локализации генов в структурах клетки различают расположенные в хромосомах ядра ядерные гены и цитоплазматические гены, локализация которых связана с хлоропластами и митохондриями.
По функциональному значению различают структурные гены, характеризующиеся уникальными последовательностями нуклеотидов, кодирующих свои белковые продукты, которые можно идентифицировать с помощью мутаций, нарушающих функцию белка, и регуляторные гены — последовательности нуклеотидов, не кодирующие специфические белки, а осуществляющие регуляцию действия гена (ингибирование, повышение активности и др.).
По влиянию на физиологические процессы в клетке различают летальные, условно летальные, супервитальные гены, гены-мутаторы, гены-антимутаторы и др.
Следует отметить, что любые биохимические и биологические процессы в организме находятся под генным контролем. Так, деление клеток (митоз, мейоз) контролируется несколькими десятками генов; группы генов осуществляют контроль восстановления генетических повреждений ДНК (репарация). Онкогены и гены — супрессоры опухолей участвуют в процессах нормального деления клеток. Индивидуальное развитие организма (онтогенез) контролируется многими сотнями генов. Мутации в генах приводят к измененному синтезу белковых продуктов и нарушению биохимических или физиологических процессов.
Гомеозисные мутации у дрозофилы позволили открыть существование генов, нормальной функцией которых является выбор или поддержание определенного пути эмбрионального развития, по которому следуют клетки. Каждый путь развития характеризуется экспрессией определенного набора генов, действие которых приводит к появлению конечного результата: глаза, голова грудь, брюшко, крыло, ноги и т. д. Исследования генов комплекса bithorax дрозофилы американским генетиком Льюисом показали, что это гигантский кластер тесно сцепленных генов, функция которых необходима для нормальной сегментации груди (thorax) и брюшка (abdomen). Подобные гены получили название гомеобоксных. Гомеобоксные гены расположены в ДНК группами и проявляют свое действие строго последовательно. Такие гены обнаружены и у млекопитающих, и они имеют высокую гомологию (сходство).
Функции генов.
В процессе реализации наследственной информации, заключенной в гене, проявляется целый ряд его свойств. Определяя возможность развития отдельного качества, присущего данной клетке или организму, ген характеризуется дискретностью действия (от лат. discretus — разделенный, прерывистый), прерывностью (интроны и экзоны). Дискретность наследственного материала, предположение о которой высказал еще Г. Мендель, подразумевает делимость его на части, являющиеся элементарными единицами, - гены. В настоящее время ген рассматривают как единицу генетической функции. Он представляет собой минимальное количество наследственного материала, которое необходимо для синтеза тРНК, рРНК или полипептида с определенными свойствами. Ген несет ответственность за формирование и передачу по наследству отдельного признака или свойства клеток, организмов данного вида. Кроме того, изменение структуры гена, возникающее в разных его участках, в конечном итоге приводит к изменению соответствующего элементарного признака.
Ввиду того что в гене заключается информация об аминокислотной последовательности определенного полипептида, его действие является специфичным. Однако в некоторых случаях одна и та же нуклеотидная последовательность может детерминировать синтез не одного, а нескольких полипептидов. Это наблюдается в случае альтернативного сплайсинга у эукариот и при перекрывании генов у фагов и прокариот. Очевидно, такую способность следует оценить как множественное, или плейотропное, действие гена (хотя традиционно под плейотропным действием гена принято понимать участие его продукта – полипептида – в разных биохимических процессах, имеющих отношение к формированию различных сложных признаков). Например, участие фермента в ускорении определенной реакции (см. рис.), которая является звеном нескольких биохимических процессов, делает зависимыми результаты этих процессов от нормального функционирования гена, кодирующего этот белок. Нарушение реакции A→B, катализируемой белком α, в результате мутации гена ведет к выключению последующих этапов формирования признаков D и E.
![]()
![]() α D
α D
![]()
![]() A B
A B
![]() C
C
E
Определяя возможность транскрибирования мРНК для синтеза конкретной полипептидной цепи, ген характеризуется дозированностью действия, т.е. количественной зависимостью результата его экспрессии от дозы соответствующего аллеля этого гена. Примером может служить зависимость степени нарушения транспортных свойств гемоглобина у человека при серповидно-клеточной анемии от дозы аллеля НЬS. Наличие в генотипе человека двойной дозы этого аллеля, приводящего к изменению структуры β-глобиновых цепей гемоглобина, сопровождается грубым нарушением формы эритроцитов и развитием клинически выраженной картины анемии вплоть до гибели. У носителей только одного аллеля НЬS при нормальном втором аллеле лишь незначительно изменяется форма эритроцитов и анемия не развивается, а организм характеризуется практически нормальной жизнеспособностью.
Современные представления о генотипе.
Геномом называют всю совокупность наследственного материала, заключенного в гаплоидном наборе хромосом клеток данного вида организмов. Геном видоспецифичен, так как представляет собой тот необходимый набор генов, который обеспечивает формирование видовых характеристик организмов в ходе их нормального онтогенеза. Например, у некоторых видов появляются гаплоидные организмы, которые развиваются на основе одинарного набора генов, заключенного в геноме. Так, у ряда видов членистоногих гаплоидными являются самцы, развивающиеся из неоплодотворенных яйцеклеток.
При половом размножении в процессе оплодотворения объединяются геномы двух родительских половых клеток, образуя генотип нового организма. Все соматические клетки такого организма обладают двойным набором генов, полученных от обоих родителей в виде определенных аллелей. Таким образом, генотип — это генетическая конституция организма, представляющая собой совокупность всех наследственных задатков его клеток, заключенных в их хромосомном наборе – кариотипе.
Кариотип – диплоидный набор хромосом, свойственный соматическим клеткам организмов данного вида, являющийся видоспецифическим признаком и характеризующийся определенным числом, строением и генетическим составом хромосом. Ниже приведены количества хромосом соматических клеток некоторых видов организмов.
Если число хромосом в гаплоидном наборе половых клеток обозначить , то общая формула кариотипа будет выглядеть как 2п, где значение различно у разных видов. Являясь видовой характеристикой организмов, кариотип может отличаться у отдельных особей некоторыми частными особенностями. Например, у представителей разного пола, имеются в основном одинаковые пары хромосом (аутосомы), но их кариотипы отличаются по одной паре хромосом (гетерохромосомы, или половые хромосомы). Иногда эти различия состоят в разном количестве гетерохромосом у самок и самцов. Чаще различия касаются строения половых хромосом, обозначаемых разными буквами – X и Y (XX или XY).
Каждый вид хромосом в кариотипе, содержащий определенный комплекс генов, представлен двумя гомологами, унаследованными от родителей с их половыми клетками. Двойной набор генов, заключенный в кариотипе,- генотип – это уникальное сочетание парных аллелей генома. В генотипе содержится программа развития конкретной особи.
Выяснилось, что множество хронических болезней человека есть проявление генетического груза, риск их развития может быть предсказан задолго до рождения ребенка на свет, и уже появились практические возможности снизить давление этого груза.
Генетический груз включает, с одной стороны, патологические генные мутации, наследуемые от родителей и прародителей, и называемые серегационным грузом, если в виде болезни проявляются рецессивные или нелетальные доминантные мутации генов (от латинского segregatio – выщепление).
С другой стороны, определенную часть этого груза составляют новые, вновь возникшие генные мутации (в результате мутагенных влияний внешней среды). Они не прослеживаются в восходящих поколениях и составляют так называемый мутационный генетический груз.
Согласно данным Н.П.Дубинина, частота спонтанных генных мутаций установлена в пределах 10-10 на геном на поколение. В геноме человека имеется около 100000 генов. Расчеты показывают, что примерно у 10% людей возникают новые мутации, вызванные мутагенным воздействием факторов окружающей среды (радиационный фон Земли, действие продуктов сжигания топлива, влияния вирусов). Безусловно, частота мутаций будет значительно выше в условиях антропогенного загрязнения внешней среды. Каждый человек наследует, как минимум, 10 скрытых мутаций, опасных для здоровья. В целом по А. Кнудсону (1986), величина постнатального генетического груза составляет 0.2 т.е. у 20% членов популяции существует вероятность развития наследственных болезней (моногенных, полигенных или связанных с мутациями генов соматических клеток).
Генетический груз проявляется, как бесплодие и спонтанные аборты, выкидыши и мертворождения, врожденные пороки и умственная отсталость. Он определяет риск гемолитической болезни новорожденных, проявления несовместимости матери и плода по ряду антигенов.
Реализация наследственной информации, заключенной в генотипе организма,— это сложный процесс, который требует тонкой регуляции для того, чтобы в клетках разной тканевой принадлежности в определенное время в процессе развития организма обеспечить синтез специфических белков в необходимом количестве.
Все клетки многоклеточного организма, возникая из зиготы путем митоза, получают полноценный набор генетической информации. Несмотря на это, они отличаются друг от друга по морфологии, биохимическим и функциональным свойствам. В основе этих различий лежит активное функционирование в разных клетках неодинаковых частей генома. Большая часть генома находится в клетках организма в неактивном, репрессированном, состоянии, и только 7—10% генов дерепрессированы, т.е. активно транскрибируются. Спектр функционирующих генов зависит от тканевой принадлежности клетки, от периода ее жизненного цикла и стадии индивидуального развития организма.
Основная масса генов, активно функционирующих в большинстве клеток организма на протяжении онтогенеза, — это гены, которые обеспечивают синтез белков общего назначения (белки рибосом, гистоны, тубулины и т.д.), тРНК и рРНК.
Фенотип – совокупность всех признаков и свойств организма, сложившихся в процессе индивидуального развития генотипа. Сюда относятся не только внешние признаки, но и внутренние: анатомические, физиологические, биохимические. Каждая особь имеет свои особенности внешнего вида, внутреннего строения, характера обмена веществ, функционирования органов, т.е. свой фенотип, который сформировался в определенных условиях среды.
Определенная роль в эволюции геномов как про-, так и эукариотических клеток принадлежит так называемым подвижным генетическим элементам — транспозонам. Они представляют собой автономные единицы, несущие в нуклеотидной последовательности информацию о структуре особых белков, которые обеспечивают их способность к перемещению из одного участка генома в другой. Такое перемещение — траспозиция — может происходить в строго определенные участки хромосом, узнаваемые этими специфическими белками. Транспозиция предполагает репликацию нуклеотидной последовательности подвижного генетического элемента и встраивание копии в ДНК с сохранением другой копии в прежнем месте.
Установлена также способность подвижных генетических элементов к точному вырезанию и удалению их из хромосомы. Перемещение таких нуклеотидных последовательностей в пределах генома может влиять на регуляцию экспрессии генов, которые прилежат к месту встраивания этих элементов. В результате таких перемещений могут активироваться ранее не активные гены, и наоборот.
Обнаружение подвижных генетических элементов в геномах как про-, так и эукариот указывает на определенные эволюционные преимущества, связанные с их наличием в наследственном материале. Возможно, рекомбинационные процессы, обеспечиваемые подвижными генетическими элементами, имеют немаловажное значение в структурной эволюции генома.
Наряду с транспозонами, не способными очевидно, существовать вне генома и образовывать свободные молекулы ДНК, описаны элементы, обнаруживаемые как в составе генома, так и вне его. Существование таких подвижных элементов дает возможность обсуждать роль горизонтального переноса генетического материала в эволюции генома.
Если описанные выше изменения структуры генома передаются из поколения в поколение организмов одного и того же вида, т.е. по вертикали, то горизонтальный перенос генетической информации может происходить и между организмами разных видов, одновременно существующими на Земле. В настоящее время доказана возможность изменения наследственных свойств у бактерий путем введения в бактериальную клетку чужеродной ДНК при конъюгации или с помощью фагов. Оказывается, чужеродную ДНК можно ввести и в эукариотическую клетку, где она будет сохраняться как внехромосомный элемент или интегрироваться в геном и экспрессироваться.
Недавно получены данные, свидетельствующие о том, что гены могут переходить от одного эукариотического организма к другому и даже от эукариот к прокариотам, хотя это происходит крайне редко. Примером могут служить данные о несовпадении скоростей эволюции отдельных последовательностей генов гистонов у некоторых видов морских ежей. Это можно объяснить относительно поздним по сравнению с временем дивергенции этих видов горизонтальным переносом указанных последовательностей, проявляющих большее сходство, чем этого можно было ожидать.
Заключение.
В заключении хотелось бы сказать о значении гена.
Ген – функциональная единица наследственности. Он играет важную роль в наследовании признаков разными организмами. На генном уровне организации наследственного материала обеспечиваются индивидуальное наследование и индивидуальное изменение отдельных признаков и свойств клеток, организмов данного вида.
Реальное существование генного уровня организации наследственного материала дало возможность исследователям при анализе характера наследования отдельных признаков открыть главные закономерности, которые легли в основу наших представлений об организации материального носителя наследственности и изменчивости. Процессы наследственности и изменчивости непосредственно влияют на ход на такой глобальный процесс как эволюция.
Мутации генов тоже оказывают большое влияние на организм.
Таким образом, значение гена велико для всех элементов нашей жизни.
Список использованной литературы.
1. Айала Ф., Кайдегер Дж.. Современная генетика: т.2. М.: Мир, 1988.
2. Биология: в 2 кн. Кн. 1: Жизнь. Гены. Клетка. Онтогенез. Человек./ под ред. В.Н.Ярыгина.
3. Биология: Большой энциклопедический словарь/ гл. ред. М.С.Гиляров.
4. Вилли К., Детье В.. Биология (биологические процессы и законы). М.: Мир, 1975.