Процесс фотосинтеза
Фотосинтез - это процесс, при котором световая энергия поглощается и используется на синтез восстановленных углеродсодержащих соединений из двуокиси углерода и воды. Этот процесс происходит только в освещенной зеленой ткани, потому что хлорофилл играет существенную роль в превращении световой энергии в химическую. Термин фотосинтез был предложен Чарльзом Рейдом Бансом из Чикагского университета в начале этого столетия. В Европе этот процесс часто называют ассимиляцией или ассимиляцией углерода. Большинство американских физиологов растений предпочитают употреблять термин ассимиляция, когда речь идет об образовании новых тканей из углеводов и азотистых соединений.
Значение фотосинтеза нельзя переоценить, потому что вся энергия, содержащаяся в нашей пище, накапливается прямо или косвенно благодаря процессу фотосинтеза, а источником большей части используемой энергии, на которой работают заводы, являются горючие ископаемые, где она была запасена посредством фотосинтеза в далеком прошлом. Лит (1972, 1975) подсчитал, что наземные растения образуют за год 100-109 т сухого вещества, из которых более 2/3 вырабатывают деревья. Общая энергия, ежегодно накапливаемая в растениях при фотосинтезе, приблизительно в 100 раз превышает энергию, содержащуюся в угле, добываемом в течение года во всех шахтах мира. Хотя леса и занимают только 1/3 поверхности суши земного шара, они вырабатывают около 2/3 сухого вещества, в то время как возделываемые земли занимают приблизительно 9% и дают только 9% сухого вещества.
Пока человек не стал широко использовать уголь, большая часть потребляемой им тепловой энергии получалась от сжигания древесины. В настоящее время древесина шире используется как строительный материал и источник получения бумаги, чем в качестве топлива, хотя интерес к последнему опять возрастает. Независимо от того, как используется древесина, необходимо помнить, что содержащиеся в ней энергия и сухое вещество накапливаются благодаря процессу фотосинтеза. В связи с этим рациональное ведение лесного хозяйства должно быть направлено на повышение количества продуктов фотосинтеза на единицу земной поверхности и эффективности их превращения в растительный материал.
Из общего количества солнечного излучения, попадающего на нашу планету, лишь половина доходит до поверхности Земли, только 1/8 имеет длину волны, подходящую для фотосинтеза, и лишь 0,4 % таких лучей (около 1 % от общего объёма энергии) используется растениями. Именно от этого одного процента зависит вся жизнь на Земле.
В процессе фотосинтеза углекислый газ в присутствии хлорофилла реагирует с водой; при этом образуется глюкоза и выделяется кислород:
6CO2 + 6H2O → C6H12O6 + 6O2.
Более грамотной будет запись
CO2 + 2H2O → (CH2O) + O2 + H2O,
Которая показывает, что выделяющийся кислород образуется из воды. Похожим уравнением описывается и хемосинтез серобактерий:
CO2 + 2H2S → (CH2O) + 2S + H2O,
 http://www.ebio.ru/images/10020201.gif
http://www.ebio.ru/images/10020201.gif
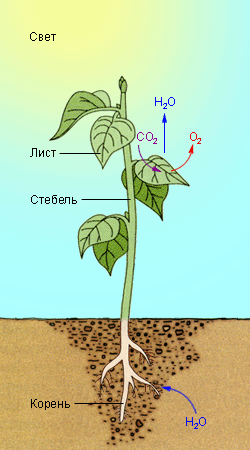
Общая схема фотосинтеза
Таким образом, процесс фотосинтеза включает в себя две стадии:
- получение водорода (фотолиз) – при этом кислород выделяется как побочный продукт реакции;
- получение глюкозы (восстановление).
 http://www.ebio.ru/images/10020202.jpg
http://www.ebio.ru/images/10020202.jpg
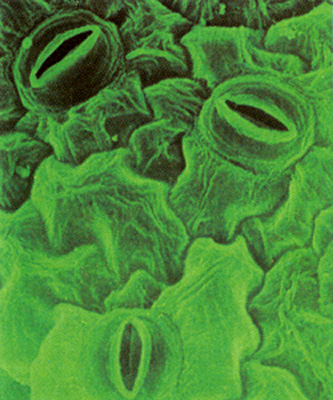
Газообмен в листе происходит через устьица
 http://www.ebio.ru/images/10020203.gif
http://www.ebio.ru/images/10020203.gif
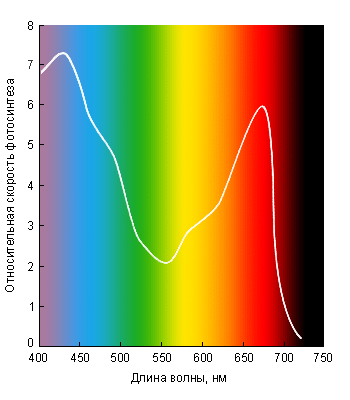
Интенсивность фотосинтеза на различных длинах волн
Первая стадия фотосинтеза протекает на свету. Световые кванты дают электронам энергию, необходимую для переноса их от хлорофилла или другого фотосинтезирующего пигмента. В ходе первой стадии из АДФ (аденозиндифосфата) и фосфата синтезируется АТФ (аденозинтрифосфат), а НАДФ (никотинамидадениндинуклеотидфосфат) восстанавливается до НАДФ∙H2. Синтез АТФ за счёт энергии световых квантов называется фотофосфорилированием. Этот процесс может быть циклическим (в реакции «работают» одни и те же электроны) и нециклическим (электроны в конце концов доходят до НАДФ и, взаимодействуя с ионами водорода, образуют НАДФ∙H2). Кислород как побочный продукт реакции выделяется только во втором случае.
Для реакций второй стадии свет не нужен. Восстановление CO2 происходит за счет энергии АТФ и накопленного НАДФ∙H2. Углекислый газ связывается с пятиуглеродным сахаром рибулозобисфосфатом, образуя две молекулы трёхуглеродной фосфоглицериновой кислоты (ФГК). Такой процесс получил название C3-фотосинтеза. Последующий цикл реакций (цикл Кальвина) приводит к образованию из ФГК сахара (например, глюкозы), а также ресинтезу рибулозобисфосфата. У некоторых растений (например, сахарного тростника, сои) наблюдается так называемый C4-фотосинтез, в реакциях которого CO2, восстанавливаясь, включается в состав органических кислот, имеющих четыре атома углерода (например, яблочной). При этом поглощение углекислоты идёт гораздо эффективнее, повышается и продуктивность растений.
На скорость фотосинтеза влияют многие факторы. Основными из них являются интенсивность света, концентрация кислорода и углекислого газа, температура окружающей среды. Состояние, когда скорость выделения кислорода растением равна скорости его дыхания, называется точкой компенсации.
Кислород в процессе фотосинтеза может действовать как конкурентный ингибитор, взаимодействуя с рибулозодисфосфатом вместо углекислого газа. При этом образуется одна молекула ФГК и фосфогликолат, сразу расщепляющийся до гликолата. Чтобы вернуть хотя бы часть углерода, связанного в бесполезном гликолате, у растения имеется процесс, называемый фотодыханием. Это зависимое от света потребление кислорода с выделением углекислого газа, заметное лишь у C3-растений, не имеет ничего общего с обычным дыханием. Фотодыхание, в целом, идёт с поглощением энергии; в результате образуется фосфоглицерат, а 25 % углерода теряется в виде CO2. В фотодыхании участвуют хлоропласты, пероксисомы и митохондрии. У C4-растений фотодыхания практически нет, что и является причиной их большей продуктивности.
В связи с энергетической проблемой учёные пытаются провести фотосинтетические процессы искусственно, особенно их первые этапы, когда вода под действием солнечной радиации расщепляется на кислород и водород. Сжигание водорода (с образованием воды) – экологически чистый процесс, который мог бы стать неплохой заменой современным источникам энергии
Хлорофилл заслуживает особого внимания, потому что в процессе фотосинтеза он является светопоглощающим пигментом, а также и потому, что создает доминирующую окраску земной поверхности. У некоторых декоративных деревьев и кустарников иногда бывают видны желтые пигменты-каротиноиды. Эти пигменты обнаруживаются и вследствие того, что условия становятся неблагоприятными для синтеза хлорофилла или его сохранения. Листья некоторых разновидностей древесных растений, например, темно-пунцовой формы бука европейского, клена дланевидного, имеют красную или пурпурную окраску из-за присутствия в клеточном соке (а не в пластидах) антоцианов. Многие другие деревья вырабатывают антоцианы осенью. (Образование осенней окраски листьев будет рассмотрено в главе 7). Хлорофилл и каротиноиды встречаются, кроме листовых пластинок, во многих тканях, в том числе в черешках, почках, семядолях, в коровой паренхиме молодых побегов и в феллодерме более старых стеблей некоторых видов. Обычно хлоропласты редко встречаются в эпидермальных клетках, за исключением замыкающих клеток устьиц. Изредка они развиваются в корнях, выставленных на свет.
Интенсивность фотосинтеза листьев древесных растений, как правило, ниже интенсивности фотосинтеза травянистых растений.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ОБРАЗОВАНИЕ ХЛОРОФИЛЛА
Пожелтение, или хлороз листьев - результат их неспособности увеличивать или сохранять содержание хлорофилла. Всесторонним изучением установлено, что это явление зависит от ряда внутренних и внешних факторов
Наиболее важный фактор - генетический потенциал растения, так как иногда в результате мутаций, приводящих к полной потере способности образовывать хлорофилл, возникают ко-роткоживущие альбиносные сеянцы. Чаще процесс синтеза хлорофилла нарушается лишь частично, проявляясь в отсутствии хлорофилла в некоторых участках листьев (пестролистность) или в общем низком содержании хлорофилла. Вследствие этого листья принимают желтоватый оттенок золотистых разновидностей деревьев и кустарников, часто используемых в качестве декоративных растений. Иногда в результате почковых мутаций возникают альбиносные или пестролистные ветви на нормальных в других отношениях деревьях и кустарниках. В синтезе хлорофилла и сборке его в фотосинтетические единицы участвуют многие гены, поэтому в образовании хлорофилла распространены аномалии. Развитие хлоропластов зависит как от ядерной, так и от плас-тидной ДНК, а также от цитоплазматических и хлоропластных рибосом.
Достаточное снабжение углеводами необходимо, по-видимому, для образования хлорофилла, и листья, испытывающие недостаток растворимых углеводов, могут не зеленеть, даже если все прочие условия благоприятные. Такие листья, опущенные в раствор сахара, обычно начинают образовывать хлорофилл. Часто образованию хлорофилла препятствуют вирусы, вызывая "желтуху", характеризующуюся общим хлорозом или бронзовой окраской листовых пластинок, а также просветлением жилок. Крапчатые листья некоторых декоративных растений - результат вирусной инфекции.
ФАКТОРЫ ВНЕШНЕЙ СРЕДЫ
Главными внешними факторами, влияющими на образование и сохранение хлорофилла, являются: свет, температура, минеральное питание, вода и кислород. Синтез хлорофилла очень чувствителен почти к любому фактору, нарушающему метаболические процессы.
Свет. Свет необходим для образования хлорофилла, хотя сеянцы некоторых хвойных и небольшое число других видов растений образуют хлорофилл в темноте. Относительно низкая освещенность эффективна для инициации или ускорения образования хлорофилла. Выращенные в темноте желтые сеянцы содержат протохлорофилл — предшественник хлорофилла а, для восстановления которого до хлорофилла а требуется свет. Очень яркий свет вызывает разложение хлорофилла. Следовательно, хлорофилл всегда одновременно синтезируется и разрушается. На ярком свету равновесие устанавливается при более низкой концентрации хлорофилла, чем при свете малой интенсивности. Теневые листья обычно имеют более высокую концентрацию хлорофилла, чем световые.
Температура. Синтез хлорофилла происходит, по-видимому, в широком интервале температур. Вечнозеленые растения умеренной зоны синтезируют хлорофилл от температур близких к температурам замерзания до самых высоких температур в середине лета. Многие хвойные становятся зимой до некоторой степени хлоротичными, вероятно, вследствие того, что распад хлорофилла превышает его синтез при очень низких температурах.
Минеральное питание. Одна из наиболее частых причин хлороза — недостаток какого-либо необходимого элемента. Недостаток азота — обычная причина хлороза древесных растений, особенно у старых листьев. Другая распространенная причина хлороза заключается в недостатке железа, преимущественно у молодых листьев. Достаточное снабжение железом необходимо, очевидно, для синтеза хлорофилла. В состав хлорофилла железо не входит, но оно служит кофактором для предшественника хлорофилла. Магний является составной частью хлорофилла, поэтому его недостаток, естественно, вызывает хлороз. Недостаток большинства макроэлементов и некоторых микроэлементов может быть причиной хлороза. Можно предположить, что почти любое нарушение нормального метаболизма препятствует синтезу хлорофилла. (Хлороз, вызываемый недостатком минеральных элементов, обсуждается более подробно в главе 10).
Вода. Умеренный водный стресс замедляет образование хлорофилла, а сильное обезвоживание растительных тканей не только нарушает синтез хлорофилла, но и вызывает распад уже имеющихся молекул. В результате листья растений, подвергшихся воздействию засухи, имеют тенденцию к пожелтению. Листья деревьев и кустарников могут также пожелтеть при насыщении водой почвы вокруг их корней. Действие засухи и плохой аэрации почвы является до некоторой степени косвенным: синтез хлорофилла задерживается вследствие общего нарушения метаболизма (см. главы 16 и 17).
Кислород. При отсутствии кислорода сеянцы не образуют хлорофилла даже на свету. Это указывает на то, что аэробное дыхание необходимо для некоторых процессов образования промежуточных соединений, что для синтеза хлорофилла необходим приток метаболической энергии.
Фотосинтез заключается в восстановлении атмосферной С02 до углеводов с использованием световой энергии, сопровождаемой освобождением кислорода из воды. Фотосинтез, подобно многим другим физиологическим процессам, состоит из нескольких последовательных этапов. Вследствие их сложности подробное обсуждение этих этапов не входит в задачу данной книги.
СВЕТОВЫЕ И ТЕМНОВЫЕ РЕАКЦИИ
Фотосинтез включает две фазы: фотохимическую, для которой требуется свет, и химическую, протекающую в темноте.
Фотохимическая фаза протекает очень быстро (за 0,00001 с). Первичная фотохимическая реакция не зависит от температуры, хотя транспорт электронов - процесс, зависящий от температуры. Темновая фаза происходит более медленно, приблизительно за 0,04 с при 25°С, замедляясь при низких температурах. Интенсивность фотосинтеза ограничивается реакцией, протекающей наиболее медленно. В связи с этим на ярком свету, когда ограничивающей является химическая фаза, процесс фотосинтеза чувствителен к температуре, а при слабой освещенности более вероятно, что ограничивающей становится фотохимическая фаза. При нормальных условиях освещения концентрация С02 чаще бывает ограничивающим фактором, чем температура, за исключением, возможно, вечнозеленых растений зимой. Большой интерес представляет поиск методов повышения эффективности фотосинтеза. Естественный отбор, действовавший в течение тысячелетий, создал согласованность биохимии и биофизики фотосинтеза, которую будет очень трудно улучшить в полевых условиях.
ПОГЛОЩЕНИЕ ДВУОКИСИ УГЛЕРОДА ФОТОСИНТЕЗИРУЮЩИМИ ТКАНЯМИ
С02 поступает преимущественно через устьица, хотя небольшое количество, по-видимому, диффундирует через кутикулу и эпидермальные клетки.
Таким образом, устьица, закрываясь, увеличивают сопротивление передвижению воды гораздо сильнее, чем сопротивление диффузии СО2, и, очевидно, понижают транспирацию в большей степени, чем фотосинтез. Это создает обоснование для использования метаболических антитранспирантов, которые вызывают закрывание устьиц, для уменьшения транспирации (см. главу 13)
ТКАНИ, ОСУЩЕСТВЛЯЮЩИЕ ФОТОСИНТЕЗ
Фотосинтез, происходящий главным образом в листьях, отмечается в некоторой степени и в других зеленых тканях, в том числе в семядолях (см. главу 15), почках, стеблях, цветках и плодах. У большинства видов фотосинтетическая деятельность тканей, не входящих в состав семядолей и листьев, незначительна. Например, фотосинтез зеленых плодов белой акации, лимона, апельсина, авокадо, винограда и сливы обычно слишком слаб для того, чтобы внести заметный вклад в их рост. Фотосинтез быстро разрастающихся зеленых шишек сосны смолистой недостаточен для того, чтобы уравновесить наблюдающееся у них интенсивное выделение СО2 в процессе дыхания. Большая часть необходимых для роста шишек углеводов мобилизуется из других источников.
Фотосинтез коры у большинства видов невелик, составляет 5% общего у тополя дельтовидного, но фотосинтез ветвей и коры очень важен для некоторых растений засушливой зоны. Ряд пустынных кустарников, сбрасывающих листья в сухое время года, имеет устойчивые фотосинтетически активные стволы и ветви. Например, Cercidium microphyllum находится в безлистном состоянии большую часть года, и его зеленые ветви могут вырабатывать до 40 % всех ежегодных продуктов фотосинтеза.
ИЗМЕНЕНИЯ ИНТЕНСИВНОСТИ ФОТОСИНТЕЗА
Интенсивность фотосинтеза древесных растений широко варьирует в зависимости от взаимодействия многих внешних и внутренних факторов, причем эти взаимодействия изменяются во времени и различны у разных видов.
Фотосинтетическую способность иногда оценивают по чистому приросту сухой массы. Такие данные имеют особое значение, потому что прирост представляет собой среднее истинное увеличение массы за большой промежуток времени в условиях внешней среды, включающих обычные периодически наступающие стрессы.
Некоторые виды покрытосеменных эффективно осуществляют фотосинтез как при низкой, так и при высокой интенсивности света. Многие голосеменные гораздо более продуктивны при высокой освещенности. Сравнение этих двух групп при низкой и высокой интенсивности света часто дает различное представление о фотосинтетической способности с точки зрения накопления питательных веществ. Кроме того, голосеменные часто накапливают некоторое количество сухой массы в период покоя, тогда как листопадные покрытосеменные теряют ее вследствие дыхания. Поэтому голосеменное растение с несколько более низкой интенсивностью фотосинтеза, чем листопадное покрытосеменное во время периода роста, может накапливать в течение года столько же или даже больше общей сухой массы благодаря гораздо большей продолжительности периода фотосинтетической активности.
ВИДОВЫЕ И ГЕНЕТИЧЕСКИЕ РАЗЛИЧИЯ
Фотосинтетическая способность часто значительно различается у разных видов, а также среди разновидностей, клонов и особей одного вида, но разного происхождения. Такое разнообразие обычно связано с основными различиями в метаболизме или в анатомической структуре листьев. Кроме того, разные виды и разновидности одного вида заметно различаются по развитию кроны, поэтому образование большего количества листьев или более продолжительный вегетационный период часто компенсируют низкую интенсивность фотосинтеза на единицу листовой поверхности или сухой массы.
Видовые различия. Некоторые исследователи сообщали о различиях в фотосинтетической активности разных видов. Например, среди лесных деревьев особенно высокая интенсивность фотосинтеза наблюдалась у некоторых покрытосеменных (у тополя, яблони, ясеня и эвкалипта) и у голосеменных (таких, как дугласия, лиственница и метасеквойя) . Обнаружено, что поглощение СО2 на единицу листовой поверхности значительно выше у дуба красного и белого, чем у дерена флоридского или сосны ладанной. Наблюдалась высокая интенсивность фотосинтеза у дугласии, средняя у сосны веймутовой и низкая у ели европейской. Определено, что метасеквойя, имеющая очень высокую интенсивность фотосинтеза и слабое дыхание, является продуктивнее бука, дуба, сосны, если или лиственницы. Часто интенсивность фотосинтеза заметно различается у видов одного рода. Например, Eucalyptus globulus имеет гораздо более высокую интенсивность, чем Е. marginata. Имеются также сообщения о различиях среди плодовых деревьев. Поглощение двуокиси углерода на единицу поверхности листьев груши и яблони в 3 раза больше, чем у листьев таких вечнозеленых многолетних растений, как цитрусовые. В других исследованиях фотосинтез изменялся в следующем порядке: яблоня > груша > вишня > слива. У листьев яблони фотосинтез был не только наиболее интенсивным, но и дольше сохранялся осенью, а слива была первой, у которой прекращалось поглощение СО2.
Генетические различия. Имеется много примеров генетических различий в интенсивности фотосинтеза. Например, обнаружены, большие различия в фото синтетической способности двух клонов лиственницы европейской. Установлено, что сеянцы дугласии по происхождению с острова Ванкувер имели при определенной температуре более высокую интенсивность фотосинтеза, чем сеянцы, происходящие из Монтаны. Однако иногда варьирование среди сеянцев одного и того же географического происхождения было большим, чем различия между сеянцами из двух источников Люк-канен и Козловский (1972) обнаружили большие различия между клонами тополя по интенсивности фотосинтеза, фотодыхания и химического (темнового) дыхания на единицу листовой поверхности и по компенсационной точке СО2, Представители секции Aigeiros имели более низкую интенсивность фотосинтеза и более высокие компенсационные точки, чем представители секции Tacamahaca.
Различия фотосинтетической способности, наблюдаемые среди генетического материала, могут быть результатом анатомических или биохимических особенностей или тех и других одновременно. Часто изменения признаков, характеризующих устьица, являются причиной различий в поглощении СО2.
Большой интерес лесных генетиков вызывает использование интенсивности фотосинтеза в качестве показателя потенциального роста деревьев. Однако между фотосинтетической способностью и ростом деревьев установлена как высокая, так и низкая и даже отрицательная корреляция. Имеются сообщения о положительной корреляции между интенсивностью фотосинтеза и ростом у тополя, у гибрида осины и тополя, у сеянцев дугласии. Некоторые исследователи обнаружили даже обратные отношения между фотосинтезом и потенциальным ростом. Например, установлено, что интенсивность фотосинтеза и рост сеянцев сосны обыкновенной зависят от происхождения семян. Однако отмечено, что у наиболее хорошо росших сеянцев была самая низкая интенсивность фотосинтеза в расчете на сухую массу листьев. У быстрорастущих сеянцев фотосинтетически активной была меньшая часть общей массы хвои. Это обусловлено более сильным взаимным затенением хвои у быстрорастущих растений или, что более вероятно, анатомическими или биохимическими различиями хвои, связанными с происхождением семян.
По мнению некоторых исследователей, кратковременные измерения фотосинтетической способности не всегда являются надежными для оценки потенциального роста, потому что его детерминируют наряду с интенсивностью фотосинтеза, по меньшей мере, еще три важные физиологические обстоятельства. Сюда входят продолжительность роста или характер сезонных изменений фотосинтеза, отношение фотосинтеза к дыханию и распределение продуктов фотосинтеза внутри дерева.
Так, у горного экотипа сосны обыкновенной с апреля по август выявлена более высокая интенсивность фотосинтеза, чем у двух долинных экотипов. Затем интенсивность фотосинтеза горного экотипа быстро уменьшалась и осенью была ниже, чем у обоих долинных экотипов. Таким образом, прогноз прироста сухого вещества по результатам измерения фотосинтеза должен основываться как на интенсивности, так и на продолжительности фотосинтеза. Важный вывод из этого исследования заключается в том, что географические расы с очень высокой интенсивностью фотосинтеза в октябре и ноябре имели и наибольшую скорость роста.
ДНЕВНОЙ ХОД ИЗМЕНЕНИЙ ФОТОСИНТЕЗА
Ранним утром светлого безоблачного теплого дня фотосинтез идет слабо вследствие небольшой интенсивности света и низкой температуры, несмотря на высокую влажность листьев и высокую концентрацию двуокиси углерода в межклетниках листьев. При повышении интенсивности света, нагревании воздуха устьица открываются, нетто-фотосинтез начинает быстро увеличиваться и может достичь максимума до полудня. Часто вслед за максимумом наступает полуденное понижение, которое может быть небольшим или резким. Полуденная депрессия нередко сменяется новым повышением фотосинтеза в более поздние послеполуденные часы, а затем окончательно снижается. Наблюдается это, как правило, ранним вечером вслед за уменьшением интенсивности света и температуры. Вследствие изменений условий внешней среды в разные дни и в пределах одного дня дневной ход фотосинтеза часто значительно отклоняется от описанного выше. Большинство дневных изменений фотосинтеза, за исключением полуденного понижения, хорошо взаимодействуют с изменениями интенсивности света.
Например, на открытом пространстве максимум интенсивности фотосинтеза трех видов покрытосеменных наблюдался в полдень и соответствовал максимуму интенсивности общей солнечной радиации. Под пологом леса интенсивность фотосинтеза значительно изменялась в течение дня. При этом наивысшая интенсивность наблюдалась в солнечных бликах. Дневной ход фотосинтеза голосеменных был очень различным в облачные и в солнечные дни. Без затенения в пасмурные или облачные дни типичная картина была такой: интенсивность нетто-фотосинтеза возрастала, достигая максимума около полудня, затем или уменьшалась, или оставалась более или менее постоянной в течение 1-2 ч и попом понижалась. В ясные солнечные дни фотосинтез обычно быстро возрастал, достигал максимума между 9 и 12 ч утра, затем, до поздних послеполуденных часов, понижался. Позднее он вновь усиливался и достигал второго максимума, но значительно более низкого.
Причины дневных изменений фотосинтеза. Регуляция дневного хода фотосинтеза под влиянием разнообразных внешних и внутренних факторов очень сложна. Главными из внешних факторов являются, по-видимому, свет, температура, наличие воды, содержание СО2 в воздухе и различные взаимодействия между ними. Внутреннюю регуляцию поглощения СО2 относили за счет разных причин: водного стресса, закрывания устьиц, чрезмерного дыхания, накопления конечных продуктов фотосинтеза и фотоокисления ферментов. Значение отдельных факторов, влияющих на поглощение СО2, нередко изменяется. Например, утром, когда клетки тургесцентны, повышение интенсивности фотосинтеза взаимосвязано с нагреванием воздуха и увеличением интенсивности света. Однако полуденный спад при наивысшей интенсивности света часто происходит, особенно в жаркие дни, вследствие слишком большой потери воды, сопровождаемой закрыванием устьиц.
Необходимо различать сезонные изменения фотосинтетической способности деревьев, вызываемые развитием листьев и состоянием метаболизма, и наблюдаемую в полевых условиях фактическую интенсивность, которую определяют как фото синтетическую способность, так и накладывающиеся друг на друга факторы внешней среды. Это разграничение важно в связи с тем, что при исследовании сезонных изменений фотосинтеза растения часто периодически переносили из открытого грунта в лабораторию и измеряли фотосинтез при стандартных и благоприятных внешних условиях. Фактическая интенсивность фотосинтеза в полевых условиях гораздо сильнее изменяется в разные дни вследствие изменений факторов внешней среды, чем интенсивность, измеренная при стандартных условиях.
Сезонные изменения фотосинтетической способности у голосеменных происходят более постепенно, чем у листопадных покрытосеменных. Когда температура весной повышается и ночные заморозки становятся менее частыми, фотосинтетическая способность голосеменных постепенно увеличивается. Осенью интенсивность фотосинтеза также постепенно понижается. У листопадных покрытосеменных фотосинтез быстро ускоряется весной, когда деревья вновь покрываются листвой, остается высоким в течение лета, быстро уменьшается в конце лета, когда листья стареют, и окончательно падает до нуля, когда они опадают. Сезонные изменения фотосинтетической способности различаются у видов с разным характером развития листьев. У видов, побеги которых полностью формируются в зимующих почках, листовая поверхность достигает максимума в начале вегетационного периода. Гетерофильные и периодически отрастающие виды продолжают увеличивать количество листьев постепенно или путем периодического образования новых побегов в течение всего сезона (см. главу 3). Следовательно, сезонные изменения фотосинтетической способности происходят при изменении площади листьев. У голосеменных фотосинтетическая способность также изменяется при появлении новой листвы, а осенью голосеменные дольше, чем покрытосеменные, сохраняют свою способность к заметному фотосинтезу. В районах с теплыми зимами фотосинтез у вечнозеленых голосеменных может происходить в течение всех месяцев года.
Общий фотосинтез дерева и характер его сезонных изменений часто заметно варьируют в разные годы вследствие различий в размерах листовой поверхности и климатических изменений.
Сезонные изменения фотосинтетической способности сеянцев сосны ладанной и сосны веймутовой в Северной Каролине исследовали Грегор и Крамер (1963). Сеянцы выдерживали на открытом воздухе. Периодически их переносили в лабораторию и измеряли поглощение СО2 при 25°С и освещенности 43000 лк. Начиная с февраля интенсивность фотосинтеза в расчете на один сеянец медленно увеличивалась у обоих видов до апреля, затем быстро возрастала и потом снижалась в течение осени и зимы. Максимальная интенсивность у сосны ладанной достигалась в середине сентября, после чего происходило быстрое осеннее понижение. Максимум фотосинтеза у сосны веймутовой наблюдался между 15 июля и 15 сентября. Осеннее снижение было более постепенным. Более высокий и оолее поздний максимум фотосинтеза сосны ладанной был обусловлен в значительной степени тем, что у сеянцев трижды отрастали побеги, прибавляя новую хвою до конца лета. В то же время сеянцы сосны веймутовой давали новые побеги только один раз, в начале вегетационного периода.
Повышение интенсивности фотосинтеза после 9 апреля отчасти могло быть отнесено за счет увеличения ассимилирующей поверхности у обоих видов. Однако значительное увеличение с 14 февраля до 9 апреля нельзя объяснить этой причиной, так как к 9 апреля новая листва еще не распустилась. Это увеличение должно было происходить в результате восстановления фотосинтетической активности уже имевшейся хвои. Уменьшение фотосинтеза после максимума, наблюдавшегося в середине сезона, было вызвано у обоих видов не потерей хвои, а скорее пониженной фотосинтетической способностью существовавшей хвои.
Сезонные изменения интенсивности фотосинтеза на единицу листвы значительно отличались от сезонного хода интенсивности фотосинтеза в пересчете на один сеянец. У сосны ладанной интенсивность фотосинтеза на единицу длины пучка хвои заметно повышалась с февраля по март, несмотря на то, что в этот период новая хвоя не появлялась. Максимальная интенсивность на единицу длины пучка была достигнута в мае, за 4 месяца до максимума интенсивности на один сеянец. Высокая интенсивность сохранялась до сентября, а затем интенсивность фотосинтеза на единицу длины хвои и на один сеянец начала понижаться. У хвои сосны веймутовой фотосинтез также заметно повышался с февраля по март, но максимум не был достигнут до июля. Высокая интенсивность сохранялась в течение сентября, несколько понизилась в ноябре и достигла минимума в январе.
Зимний фотосинтез вечнозеленых растений. В районах с мягкими зимами у вечнозеленых растений фотосинтез осуществляется в течение всего года. В местах с суровыми зимами фотосинтез может быть незначительным несколько недель или месяцев.
Наблюдаемое поглощение СО2 или прирост сухой массы в течение зимы были установлены для сосны ежовой и сосны ладанной на юге Соединенных Штатов, для сосны замечательной и сосны обыкновенной в Аберистуите (Уэльс) и для широколистных вечнозеленых растений в Северной Италии. Деревья дугласии накапливали значительные количества продуктов фотосинтеза зимой в мягких климатических условиях прибрежного района северо-запада Соединенных Штатов. В засушливый год чистый прирост продуктов фотосинтеза в течение зимы приближался к 1/4 общего прироста за весь год. Вдоль всей прибрежной зоны Норвегии у сосен и елей уменьшалась сухая масса в течение коротких периодов зимой, но за всю зиму у них обнаружился чистый прирост, доказавший превышение фотосинтеза над дыханием. Сухая масса сеянцев ели ситхинской в Южной Шотландии удваивалась за период с конца сентября и до середины апреля. Большая часть прироста приходилась на конец марта - начало апреля, но некоторое количество сухой массы накапливалось в середине зимы.
В районах с холодными зимами интенсивность фотосинтеза у вечнозеленых растений наблюдалась вплоть до точки замерзания или даже немного ниже. Например, около Мюнхена поглощение СО2 елью европейской происходило в дни, когда температура воздуха была немного ниже нуля. В долине около Инсбрука (Австрия) фотосинтез ели европейской продолжался и зимой до наступления сильных морозов. Однако на границе древесной растительности около горы Патчеркофель морозная погода в ноябре (от -10° до -15°) вызвала прекращение фотосинтеза. Пизек и Випклер (1958) показали, что интенсивность нетто-фотосинтеза ели европейской и сосны кедровой европейской была заметной до поздней осени. Последующие изменения температуры на несколько градусов ниже и выше нуля вызывали колебания поглощения СО2. Как только температура падала ниже -4° или -5°С, фотосинтез прекращался. Если же после этого морозы повторялись в течение нескольких ночей, то днем фотосинтез был ингибирован, даже если температура поднималась выше нуля. После мороза от -6° до -8°С нетто-фотосинтез прекращался, и требовалось несколько дней с мягкой погодой для восстановления фотосинтетической способности. Полностью фотосинтез возобновлялся с повышением температуры весной. Особенно это касалось частей кроны, в которых происходил распад хлорофилла в течение зимы. Весной вместе с колебаниями температуры колебалась и интенсивность фотосинтеза. Следовательно, фотосинтетический аппарат оставался функционально активным только до тех пор, пока зимой не наступали морозы, У верхней границы распространения леса температуры были в течение 4-5 месяцев такими низкими, что фотосинтез по существу был невозможен.