Природа сна
Что такое сон, для чего он нужен организму? Вопрос о функциональном назначении столь обыденного состояния кажется настолько наивным, что даже не требует раздумий: конечно, для отдыха! Однако такой ответ порождает цепочку новых вопросов: что такое отдых? Почему он столь продолжителен и столь сложно организован? Почему приурочен к определенным периодам суток? Почему для отдыха недостаточно телесного покоя, а необходимо еще и выключение органов чувств, что, казалось бы, резко повышает уязвимость по отношению к неблагоприятным факторам среды? Почему теплокровные животные, у которых “постоянство внутренней среды является залогом свободной жизни”, вынуждены, подобно своим холоднокровным предкам, каждые сутки на несколько часов впадать в состояние неподвижности и ареактивности?
На протяжении многих столетий сон рассматривался именно по этим внешним признакам, т.е. состоянию покоя и пониженной реактивности. Такому подходу не смогло помешать даже формирование представлений о двух состояниях “внутри” естественного сна, принципиально отличных друг от друга и от бодрствования (медленноволновая и парадоксальная фазы).
Однако в последнее время появляется все большее число фактов, которые не укладываются в такие представления. Так, в начале 80-х годов сотрудники I Московского медицинского института В.С.Ротенберг и С.И.Кобрин, изучая сон больных с полной атрофией мышечной системы, не выявили его сокращения, хотя эти больные вовсе не нуждались в соматическом (телесном) “отдыхе”. Значит, сон не есть покой, а телесный отдых вовсе не обязательный элемент физиологического сна.
Аналогичным образом можно рассмотреть и такую общепринятую характеристику сна, как ареактивность, т.е. психическую заторможенность, отсутствие реакции на внешние стимулы. Во-первых, это “апостериорный” признак сна, поскольку порог пробуждения можно определить, лишь разбудив человека. Во-вторых, ареактивность, так же как и неподвижность, не служит достаточным признаком, поскольку она характерна для целого ряда заболеваний и других патологических состояний: фармакологического сна, наркоза, комы и пр.
Какой сон важнее - медленный или быстрый?
Появление электроэнцефалографии во второй половине XX в. позволило, наконец, строго разграничить фазы сна и тем самым подойти к выяснению их физиологической роли. Поскольку идентификацию сна, его фаз и стадий физиологи проводят на основе общепринятых, так называемых полиграфических, критериев, полиграмм - электроэнцефалограммы (ЭЭГ), электромиограммы (ЭМГ), электроокулограммы (ЭОГ), то естественно определять сущность сна по этим показателям. Однако и здесь мы сталкиваемся с теми же трудностями: нет ни одного признака, достаточного для определения сна. Отдельные характеристики медленного и парадоксального сна на ЭЭГ иногда встречаются и в других состояниях. Так, при различных формах патологии и под влиянием фармакологических препаратов на ЭЭГ наблюдаются те или иные изменения, “имитирующие” определенные стадии сна.
Ночной сон человека. Видно, что в течение ночи проходят четыре цикла сна, причем они неравнозначны: в первую половину ночи преобладает глубокий медленный сон, а в утренние часы - парадоксальный, сопровождаемый сновидениями. Смена поз характерна до и после каждого периода парадоксального сна (выделены цветом), который сопровождается интенсивной психической активностью, движением глаз и активностью нейронов ретикулярной формации ствола.
Вероятнее всего необходимым и достаточным признаком сна можно считать ритмичность, т.е. чередование определенных физиологических признаков (полиграфических картин), позволяющих отличить нормальный сон от монотонных “сноподобных состояний”. Соответственно и критерием “нормальности” сна служит циклическое чередование стадий 1-2-3-4 медленного сна, которое завершается парадоксальной фазой. На основе такого подхода современное определение сна звучит следующим образом: это “особое генетически детерминированное состояние организма человека (и теплокровных животных, т.е. млекопитающих и птиц), характеризующееся закономерной последовательной сменой определенных полиграфических картин в виде циклов, фаз и стадий” (1).
Что же лежит за этим циклическим чередованием? Каково назначение каждой из двух фаз сна? В физиологии для понимания функций отдельного органа применяют классический метод разрушения: если повредить или удалить данный орган, то, зная последствия и адекватно истолковывая их, можно выяснить его роль. Подобный подход используют и в отношении сна: не давать испытуемому или подопытному животному спать в течение некоторого времени и посмотреть, что при этом меняется в организме и поведении. Впервые такие опыты выполнила более 100 лет назад русская ученая М.М.Манасеина (1843-1903), которая стала в сущности основоположником “науки о сне” - сомнологии.
В нашем столетии в экспериментах на животных и в наблюдениях за здоровыми людьми неоднократно пытались выяснить, к чему приводит лишение сна (депривация). Однако лишь с использованием электроэнцефалографии такие попытки получили научное обоснование. Исследования последних лет на людях дали до некоторой степени парадоксальные результаты: депривация в течение одних или нескольких суток наиболее мягким, щадящим способом не приводила к серьезным нарушениям в организме и психике субъектов. Наблюдалась лишь повышенная сонливость, утомляемость, раздражительность, рассеянность. Казалось, что главный результат лишения сна - нарастающая потребность в нем!
Естественно, подобные работы на людях не могут длиться более 2-3 суток; поэтому последствия длительного лишения сна изучают только в опытах на животных. Так, в 80-е годы группа американских специалистов (А.Речшаффен и сотрудники) получила принципиально важные результаты. Как показали эксперименты, если при первых признаках сна на ЭЭГ (появление сонных веретен и дельта-волн) животных будить, то наступает временное “дробление” сна на очень короткие периоды и его пространственная “локализация”, когда сон протекает в отдельных участках мозга (2). Подобный феномен в опытах на обезьянах описал И.Н.Пигарев (3) (Институт проблем передачи информации РАН), а Л.М.Мухаметов с сотрудниками (Институт проблем экологии и эволюции им.А.Н.Северцова РАН) наблюдали чередующийся однополушарный медленный сон у дельфинов и ушастых тюленей (4).
Сопоставив эти результаты с некоторыми другими данными по хронической депривации с помощью физических методов, мы пришли к неожиданному выводу: полностью исключить медленный сон в принципе невозможно.
Как показали эксперименты, через несколько недель от начала хронической депривации у крыс “давление” медленного сна уменьшилось, и если депривация прекращалась, то “отдачи” медленного сна не наблюдалось. Ясно, что вначале это “давление” растет, а затем, по достижении некоторого критического уровня, - спадает на нет в результате постепенной адаптации феноменов и структуры медленного сна к условиям депривации.
Совершенно противоположные результаты получены в отношении парадоксального сна. Опыты Речшаффена и сотрудников продемонстрировали, что, какой бы вид депривации сна ни проводился (тотальное лишение сна, избирательное лишение медленной или парадоксальной фазы), в результате всегда критично угнетение именно парадоксального сна. Рано или поздно оно приводит к одним и тем же драматическим последствиям (изменению внешнего вида, поведения и внутренних органов), которые через несколько “бессонных” недель завершаются неизбежной гибелью животных. Характерно, однако, что непосредственной причины их гибели обнаружить не удалось.
Интересно, что у крыс наблюдалось резкое падение амплитуды ЭЭГ после хронической депривации, которое возникало каждый раз примерно за сутки до гибели животного. Если на этом фоне эксперимент прекращался, то крыса уже не могла заснуть и амплитуда ЭЭГ не восстанавливалась; смерть все равно наступала в течение суток. Следовательно, это падение амплитуды ЭЭГ указывало на какое-то необратимое нарушение работы мозга, вызванное лишением парадоксального сна. Если же опыт прекращался на позднем этапе депривации, но до наступления этого критического момента, то наблюдалась мощная “отдача” только парадоксального сна, независимо от того, какой вид депривации применялся - лишение всего сна, парадоксального или медленного.
Таким образом, опыты с длительным лишением сна у лабораторных животных еще раз показывают, что сон включает два принципиально различных состояния организма - медленноволновую и парадоксальную (быструю) фазы, подтверждая гениальную догадку М.Жуве, впервые высказанную почти 40 лет назад.
Механизмы сна
Один из главных вопросов, волновавших физиологов еще со времен Павлова, - это существование в мозге “центра сна”. Во второй половине нашего столетия прямое изучение нейронов, вовлеченных в регуляцию сна-бодрствования, показало, что нормальная работа таламо-кортикальной системы мозга, обеспечивающая сознательную деятельность человека в бодрствовании, возможна только при участии определенных подкорковых, так называемых активирующих, структур (5). Благодаря их действиям в бодрствовании мембрана большинства кортикальных нейронов деполяризована на 10-15 мВ по сравнению с потенциалом покоя – (65-70) мВ. Только в состоянии этой тонической деполяризации нейроны способны обрабатывать информацию и отвечать на сигналы, приходящие к ним от других нервных клеток (рецепторных и внутримозговых).
Как сейчас ясно, таких систем тонической деполяризации, или активации мозга (условно “центров бодрствования”), несколько - вероятно, пять или шесть. Располагаются они на всех уровнях мозговой оси: в ретикулярной формации ствола, в области голубого пятна и дорзальных ядер шва, в заднем гипоталамусе и базальных ядрах переднего мозга. Нейроны этих отделов выделяют медиаторы - глутаминовую и аспарагиновую кислоты, ацетилхолин, норадреналин, серотонин и гистамин, активность которых регулируют многочисленные пептиды, находящиеся с ними в одних и тех же везикулах. У человека нарушение деятельности любой из этих систем не компенсируется за счет других, несовместимо с сознанием и приводит к коме.
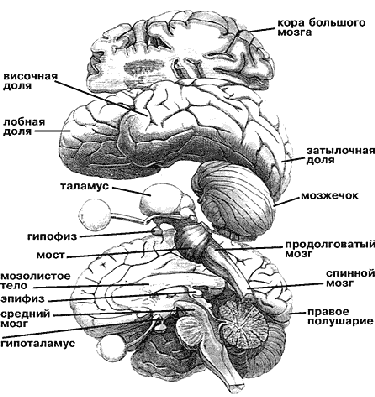
Общая схема строения мозга и его фрагмент (внизу), где в основном расположены центры бодрствования и парадоксального сна (выделены цветом); указаны соответствующие медиаторы.
Казалось бы, если в мозге есть “центры бодрствования”, то по крайней мере должен быть один “центр сна”. Однако в последние годы выяснилось, что в сами “центры бодрствования” встроен механизм положительной обратной связи. Это особые нейроны, которые осуществляют торможение активирующих нейронов и сами тормозятся ими. Такие нейроны разбросаны по разным отделам мозга, хотя больше всего их в ретикулярной части черного вещества. Все они выделяют один и тот же медиатор - гамма-аминомасляную кислоту, главное тормозное вещество мозга. Стоит только активирующим нейронам ослабить свою деятельность, как включаются тормозные нейроны и ослабляют ее еще сильнее. В течение некоторого времени процесс развивается по нисходящей, пока не срабатывает некий “триггер” и вся система переключается либо в состояние бодрствования, либо парадоксального сна. Объективно этот процесс отражает смена картин электрической активности головного мозга (ЭЭГ) по ходу одного полного цикла сна человека (90 мин).
В последнее время внимание исследователей привлечено еще к одной эволюционно древней тормозной системе головного мозга, использующей в качестве медиатора нуклеозид аденозин. Японский физиолог О.Хаяйси с коллегами показали, что синтезируемый в мозге простагландин D2 участвует в модуляции аденозинэргических нейронов. Поскольку главный фермент этой системы - простагландиназа-D - локализован в мозговых оболочках и хороидном плексусе, очевидна роль этих структур в формировании определенных видов патологии сна: гиперсомнии при некоторых черепно-мозговых травмах и воспалительных процессах менингеальных оболочек, африканской “сонной болезни”, вызываемой трипаносомой, которая передается через укусы мухи цеце и пр.
Прямая регистрация одиночной активности нейронов мозга в экспериментах на лабораторных 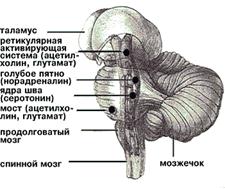 животных показала, что в бодрствовании (в состоянии тонической деполяризации) характер разрядов таламо-кортикальных клеток высоко индивидуален. Но по мере углубления сна и нарастания синхронизированной активности в ЭЭГ начинают преобладать более мощные тормозные постсинаптические потенциалы, перемежающиеся периодами экзальтации - высокочастотными вспышками нейронных разрядов (такой рисунок нейронной активности называется “пачка-пауза”).
животных показала, что в бодрствовании (в состоянии тонической деполяризации) характер разрядов таламо-кортикальных клеток высоко индивидуален. Но по мере углубления сна и нарастания синхронизированной активности в ЭЭГ начинают преобладать более мощные тормозные постсинаптические потенциалы, перемежающиеся периодами экзальтации - высокочастотными вспышками нейронных разрядов (такой рисунок нейронной активности называется “пачка-пауза”).
Тогда появляется “хоровая” активность нейронов, и условия для переработки информации в мозге, причем не только поступающей от органов чувств, но и хранящейся в памяти, резко ухудшаются. Однако средняя частота импульсации корковых и таламических нейронов не снижается, а у ГАМК-эргических (тормозных) нейронов она даже значительно повышается. Что касается активирующих нейронов, то их разряды становятся реже. Эти нейрофизиологические феномены хорошо коррелируют с известными данными о постепенном торможении психической активности по мере углубления медленного сна у человека (6).
Если с точки зрения нейронной активности бодрствование - это состояние тонической деполяризации, то медленный сон - тоническая гиперполяризация. При этом направление движения через клеточную мембрану основных ионных потоков (катионов Na+, K+, Ca2+, анионов Cl–), а также важнейших макромолекул меняется на противоположное.
Таким образом, можно было бы сказать, что во время медленного сна восстанавливается мозговой гомеостаз, нарушенный в ходе многочасового бодрствования. С этой точки зрения бодрствование и медленный сон - как бы “две стороны одной медали”. Периоды тонической деполяризации и гиперполяризации должны периодически сменять друг друга, чтобы сохранить постоянство внутренней среды головного мозга и обеспечить нормальную работу таламо-кортикальной системы - субстрата высших психических функций человека. Отсюда ясно, почему в мозге нет единого “центра медленного сна” - это значительно уменьшило бы надежность всей системы, сделало бы ее более жестко детерминированной, полностью зависящей от “капризов” этого центра в случае каких-либо нарушений его работы.
С другой стороны, становится также понятно, почему почти невозможно длительное полное подавление медленного сна: в норме активность периодически сменяется покоем, бодрствование - медленным сном, охватывающим весь мозг целиком. Известно, что при искусственной хронической депривации механизмы бодрствования и медленного сна начинают функционировать диффузно и одновременно. При этом, разумеется, страдает нормальное поведение, зато, несмотря на депривирующее воздействие, восстанавливается мозговой гомеостаз.
Однако и здесь все не так просто. Недавно Пигарев в опытах на кошках показал, что по мере развития синхронизации в ЭЭГ первичные нейроны зрительной и слуховой коры перестают реагировать на специфические стимулы и начинают все в большей степени отвечать на импульсацию, приходящую в кору со стороны внутренних органов (7). Принимая во внимание обнаруженные особые Ca–-каналы на мембране многих корковых нейронов, которые открываются при гиперполяризации, можно предположить, что в медленном сне мозг не прекращает перерабатывать информацию, а переходит от обработки внешних сигналов к интероцептивной импульсации.
Таким образом, на пороге XXI в. функция медленного сна, кажется, начинает, наконец, вырисовываться: это восстановление гомеостаза мозговой ткани и оптимизация управления внутренними органами. Для гигиены сна это означает подтверждение старого, как мир, но почему-то забытого правила: без хорошего сна не может быть хорошего бодрствования!
Совершенно по-другому обстоит дело с парадоксальным сном, который, в отличие от медленного сна, имеет ярко выраженную активную природу (8). Парадоксальный сон запускается из четко очерченного центра, расположенного в задней части мозга, в области варолиева моста и продолговатого мозга, а медиаторами служат ацетилхолин, глутаминовая и аспарагиновая кислоты. Во время парадоксального сна клетки мозга чрезвычайно активны, но информация от органов чувств к ним не поступает и не подается на мышечную систему. В этом и заключается парадоксальность этого состояния (9).
Фрагменты полиграммы на разных стадиях сна. Видно, что для смены стадий медленного сна характерно постепенное увеличение амплитуды и снижение частоты волн ЭЭГ, смена быстрых движений глаз медленными, вплоть до полного исчезновения (ЭОГ регистрируется на фоне ЭЭГ и выделены цветом), прогрессивное уменьшение амплитуды ЭМГ. При парадоксальном сне ЭЭГ такая же, как при бодрствовании, ЭОГ демонстрирует быстрые движения глаз, а ЭМГ почти не регистрируется.
Видимо, при этом интенсивно перерабатывается информация, полученная в предшествующем бодрствовании и хранящаяся в памяти. Согласно гипотезе Жуве, в парадоксальном сне, пока непонятно как, в нейрологическую память передается наследственная, генетическая информация, имеющая отношение к организации целостного поведения. Подтверждением таких психических процессов служит появление в парадоксальном сне эмоционально окрашенных сновидений у человека, а также обнаруженный Жуве с сотрудниками и детально исследованный Э.Моррисоном с коллегами феномен демонстрации сновидений у подопытных кошек (10). Они выяснили, что в мозге кошек имеется особая область, ответственная за мышечный паралич во время парадоксального сна. Если ее разрушить, подопытные кошки начинают показывать свой сон: убегать от воображаемой собаки, ловить воображаемую мышь и т.д. Интересно, что “эротические” сны у кошек никогда не наблюдались, даже в брачный сезон.
Хотя в парадоксальном сне некоторые нейроны ретикулярной формации ствола и таламо-кортикальной системы демонстрируют своеобразный рисунок активности, различия между мозговой деятельностью в бодрствовании и парадоксальном сне довольно долго выявить не удавалось. Это было сделано лишь в 80-е годы. Оказалось, что из всех известных активирующих мозговых систем, которые включаются при пробуждении и действуют во время бодрствования, в парадоксальном сне активны лишь одна-две. Это системы, расположенные в ретикулярной формации ствола и базальных ядрах переднего мозга, использующие в качестве передатчиков ацетилхолин, глутаминовую и аспарагиновую кислоты. Все же остальные активирующие медиаторы (норадреналин, серотонин и гистамин) в парадоксальном сне не работают. Это молчание моноаминоэргических нейронов ствола мозга определяет различие между бодрствованием и парадоксальным сном, или на психическом уровне - различие между восприятием внешнего мира и сновидений (11).
Непонятным оставалось все же, как эта активация, столь отличная от бодрствования, отражается на работе коры. Лишь в 1996-1997 гг. три независимых исследования выявили в парадоксальном сне (методом позитронной эмиссионной томографии) высокоспецифичный характер пространственного распределения активации и инактивации определенных участков коры и некоторых подкорковых ядер в мозге человека (12).
Павлов и природа сна
Известно, что Павлов чрезвычайно интересовался проблемой сна и считал ее одной из ключевых в изучении высшей нервной деятельности. Все знают его определение сна как “разлитого коркового торможения”. После открытия парадоксального сна казалось, что павловская теория в этой своей части безнадежно устарела. Разумеется, мысль о создании физиологии сновидений “и во сне не могла ему присниться”, если уместно привести такой каламбур. Однако, справедливости ради, следует напомнить, что идея о трех формах существования - бодрствование, спокойный сон и сон со сновидениями - впервые прозвучала в Упанишадах, древнеиндийском эпосе. В истории же европейской культуры такое, кажется, не приходило в голову никогда и никому до Жуве. Даже первооткрыватели парадоксального сна - Н.Клейтман, Ю.Азеринский и В.Демент назвали это состояние stage-1-REM, т.е. стадией засыпания (дремотой) с быстрыми движениями глаз, воспринимая его лишь как переходное между бодрствованием и сном!
Если же взять медленноволновый (ортодоксальный) сон, сон вообще, то сейчас, на рубеже веков, уместно задаться вопросом: а так ли уж неправ был Павлов в своих представлениях о сне? Разумеется, в ту, “доэлектрофизиологическую” эпоху эти представления могли быть лишь чисто интуитивными. Но читатель настоящей статьи, зная о мощной активации тормозных нейронов и выбросе их медиаторов - ГАМК и аденозина в медленном сне, об активации, которая начинается в локальных таламо-кортикальных областях и постепенно распространяется по всей системе, о тонической гиперполяризации как периоде своеобразного функционального восстановления нейронов и т.п., вправе сам судить, подвела ли интуиция на сей раз гениального ученого.
В конце своей долгой жизни, в 1935 г., Павлов высказал следующую мысль: “Ясное дело, что наша дневная работа представляет сумму раздражений, которая обуславливает известную сумму истощения, и тогда эта сумма истощения, дошедшая до конца, и вызывает автоматически, внутренним гуморальным путем , тормозное состояние, сопровождаемое сном”. Эту формулировку можно назвать пророческой - она звучит вполне актуально и в наши дни.
1. Ковальзон В.М. // Журн. эволюц. биохимии и физиологии. 1993. Т.29. № 5-6. С.627-634.
2. Rechtschaffen A. et al. // Science. 1983. V.221. № 4606. P.182; Sleep Research. 1984. V.13. P.185, 190, 191.
3. Pigarev I.N. // Neuroreport. 1997. V.8. № 11. P.2557-2560.
4. Ковальзон В.М., Мухаметов Л.М. // Журн. эволюц. биохимии и физиологии. 1982. Т.18. № 3. С.307-310.
5. Steriade M. // Science. 1996. V.272. P.225-226.
6. Steriade M. // Neurology. 1992. V.42. Suppl.6. P.9-18.
7. Pigarev I.N. // Neurosсience. 1994. V.62. P.1237-1243.
8. Jones B.E. // Neuroscience. 1991. V.40. № 3. P.637-656.
9. Jouvet M. // Sleep. 1994. V.17. № 8. P.S77-S83.
10. Моррисон Э. В мире науки. 1983. № 6. С.62-71.
11. Hobson J.A., Stickgold R., P a c e - S c h o t t E.F. // Neuroreport. 1998. V.9. № 3. P.R1-R14.
12. Maquet P., Peters F.-M., Aerts J. et al. // Nature. 1996. V. 383. P. 163-166.
13. В.М.Ковальзон, доктор биологических наук. Институт проблем экологии и эволюции им. А.Н.Северцова РАН, Москва. "Природа сна"