Клещи рода dermacentor koch. 1844 в Крыму
Евстафьев И.Л.
Среди всего разнообразия видов - консументов второго порядка, особое место занимает древняя и трофически высокоспециализированная группа эктопаразитов - иксодовых клещей (сем. Ixodidae)
Иксодовые клещи - один из важнейших компонентов паразитокомплексов млекопитающих Крымского полуострова. Фауна клещей Крыма изучена достаточно полно, однако по их экологии имеются только отрывочные сведения более чем тридцатилетней давности (Вшивков, 1957, 1958, Емчук, 1957, Мельникова, 1963, 1961, 1955). Данная работа посвящена экологии клещей рода Dermacentor Koch., повсеместно распространенных в пределах полуострова и имеющих важное эпизиоотическое значение в циркуляции возбудителей ряда природно-очаговых инфекций (Гутова, Сиротюк, 1989, Маркешин, Караванов, Ковин, Захарова, Евстратов, Бычкова, Евстафьев. 1989, Маркешин, Смирнова, Евстафьев,1992, Маркешин , 1994)
Сбор иксодовых клещей проводился по общепринятым методикам на стандартный флаг, и вручную - с различных домашних животных (собак, крупного и мелкого рогатого скота (Фасулати, 1971). Клещи очесывались с мышевидных грызунов и землероек, добытых на давилки Геро при зоопаразитологическом обследовании территории полуострова.
Ареалы клещи рода Dermacentor и их стациальное распределение
Род Dermacentor представлен на Крымском полуострове двумя видами: D. marginatus Sulz. и D. reticulatus Koch. За 1986-1997 годы на территории полуострова поймано 4881 экз. клещей р. Dermacentor: 62,7 % составили клещи D.marginatus, 37,3 % - D. reticulatus.
Ареал D. marginatus занимает практически весь полуостров, за исключением яйл (безлесных высокогорных крымских лугов) и лесных массивов, примыкающих к яйлам. Ареал D. reticulatus образует в Крыму географически изолированную от основного ареала популяцию, ограниченную горно-лесной зоной (Рис. 1).
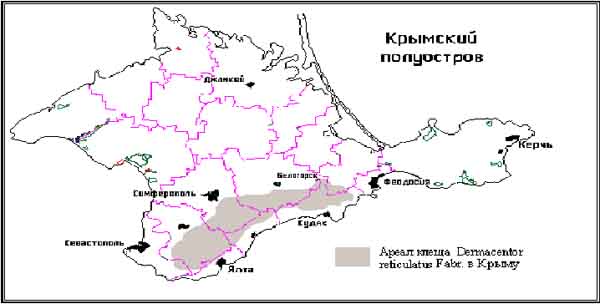
Рис. 1.
К характерным морфологическим признакам рода Dermacentor относится наличие светлых эмалевых пигментов в виде пятен различной формы и размеров, лучше всего выраженных на спинном щитке, и в меньшей мере - на ногах и хоботке. Форма эмалевых пятен и их количество весьма значительно варьирует в пределах одного вида и даже одной популяции (Померанцев Б.И., 1950, Померанцев, Матикашвили, 1940, Емчук Е.М, 1960). У разных видов рода имеются опре-деленные закономерности в развитии этого пигмента. D. daghestanicus - житель наиболее засушливых мест, почти совер-шенно белый. D. reticulatus и D. marginatus, населяющие местности с относительно большей влажностью, окрашены темнее. Можно предположить, что белый пигмент спинного щитка, характерный для рода Dermacentor, также является следствием происхождения этих клещей из мест, для которых характерны пониженная влажность воздуха и повышенная инсоляция. Многие факты подтверждают предположение о существовании связи между окраской клещей и количеством осадков. Большое значение имеют микроклиматы тех мест, где проходят свое разви-тие молодые фазы клещей, в частности их нимфы (Резник, 1970).
Виды рода Dermacentor характеризуются вполне определенными требованиями к гидротермическим условиям среды обитания, что проявляется в при-уроченности к определенным природно-климатическим зонам и типам биотопов. Распределение клещей в пределах своего ареала на территории полуострова крайне неравномерное и носит ярко выраженный агрегированный характер. Клещи образуют локальные очажки с более высокой численностью в стациях, характеризующихся оптимальными для вида экологическими условиями, к которым приспособился данный вид в процессе филогенетического развития.
В Крыму по комплексу характеристик, включающих особенности климата, ландшафта, растительности и др., наиболее естественно выделение трех крупных ландшафтно-географических единиц: горно-лесной и равнинно-степной зон и территории Керченского полуострова (Кочкин, 1967, Геоботанiчне районування:, 1977).
По гидротермическим условиям для большинства видов клещей с пастбищным типом паразитирования наиболее благоприятны лесистая территория горного и предгорного Крыма, которая занимает около 20% площади полуострова и характеризуется наличием множества разнообразных природных биотопов, образованных как травянистой, так и древесно-кустарниковой растительностью, со всеми промежуточными типами. Для данной зоны характерна повышенная влажность (выпадает 600-1000 мм осадков), что благоприятствует длительному выживанию клещей (Кочкин, 1967).
Равнинно-степной Крым занимает около 70% площади Крымского полуострова. Отличительной чертой этого района является плоский рельеф, господство культурного ландшафта, малое количество осадков, низкая относительная влажность воздуха, что в целом создает неблагоприятные условия для клещей (Геоботанiчне районування, 1977).
Промежуточное положение по условиям существования для клещей занимает Керченский полуостров, территория которого составляет около 1/10 площади полуострова. Значительная пересеченность рельефа благоприятствует сохранению здесь больших нераспаханных участков, которые используются для выпаса скота, что создает благоприятные условия для развития иксодовых клещей. Район характеризуется засушливостью (годовая сумма осадков колеблется в пределах 270-400 мм), однако близость моря благотворно сказывается на влажности воздуха, которая никогда не бывает такой низкой, как в равнинно-степном Крыму. Отмеченные выше закономерности обуславливают формирование своеобразных комплексов клещей в отмеченных регионах.
Несмотря на то, что Крымские горы имеют относительно небольшие высоты (до 1500 м над уровнем моря), выявлены определенные высотные закономерности в распространении клещей. Так, в степных районах, вплоть до предгорий, встречается только D. marginatus. Этот же вид повсеместно доминирует и в предгорных районах, хотя, начиная с высот 250-300 м над уровнем моря в наиболее влажных и затененных стациях древесно-кустарниковых сообществ, начинают встречаться клещи D. reticulatus. В среднем поясе гор во многих стациях оба вида встречаются совместно, при незначительном доминировании одного либо другого вида, что обусловлено особенностями стации. В высокогорных районах (яйлах и примыкающих лесных массивах) доминируют клещи D. reticulatus, находки клещей D. marginatus здесь случайны и единичны. По лесным массивам клещи D. reticulatus проникают и на Южный Берег, при этом не встречаясь на высотах ниже 500-700 м. Здесь же, на ЮБК, в нижнем поясе гор (вплоть до морского побережья), встречаются D. marginatus, хотя численность их мала..
D. marginatus - лесостепной вид, характеризуется теплолюбивостью и достаточно высокой устойчивостью к сухости (мезофил-ксерофил). Поэтому он встречается практически во всех зонах полуострова, однако основная часть популяции сконцентрирована в предгорьях и нижнем поясе Крымских гор, встречаясь на опушках, лесных полянах, вырубках, избегая сплошных лесных массивов. В степных районах D. marginatus встречается в искусственных лесонасаждения (Марьевское лесничество Керченского п-ва, лесопосадки в р-не п. Щелкино и др.), лесополосах, по балкам и на целинных участках, заросших кустарниками (терн, шиповник и др.) и травянистой растительностью. В открытых стациях, лишенных древесно-кустаниковых насаждений вид крайне редок, куда, по-видимому, попадает случайно - с прокормителями.
D. reticulatus - типичный обитатель зоны лиственных и смешанных лесов. В высокоствольных лесах, лишенных травянистой растительности, он не встречается, а приурочен к полянам, вырубкам, опушкам и горным лугам яйл. D. reticulatus отмечается в стациях, расположенных на высотах с 300-400 м над у.м. и выше, встречаясь до 1100 м (Ай-Петринская яйла) и выше.
Аналогичное стациальное размещение отмечалось авторами и в других частях ареала этих видов (Олсуфьев, 1940, Галузо, 1948, Федюшин, 1960). Нельзя только согласиться с утверждением (Резник, 1970), что клещи рода Dermacentor связаны в биотопами, отличающимися меньшей степенью увлажнения и более высоким прогреванием по сравнению с остальными биотопами той природной зоны, где они встречаются, так как в Крыму отмечается обратная зависимость.
Таким образом, в условиях сухого Крымского климата, оба вида характеризуются определенной гигрофильностью. Они избегают ксерофильных участков с постоянно высокой степенью инсоляции, занимая экотонные сообщества (на границе древесной и травянистой растительности) с умеренной влажностью почвы и подстилки.
Прокормители (хозяева) клещей рода Dermacentor
Клещи рода Dermacentor отличаются большим однообразием в выборе хозяев, как для взрослой, так и для предимагинальных фаз развития. Личинки и нимфы почти исключительно питаются на мелких млекопитающих (представителях отрядов грызунов и насекомоядных), но иногда встречаются и на птицах (Резник, 1970). На стадии имаго клещи трофически связаны с крупными и средними млекопитающими.
Видовой состав мелких млекопитающих - прокормителей предимагинальных стадий клещей, в горно-лесных и равнинных степных районах существенно отличается, как по видовому, так и по численному составу. В горно-лесной зоне доминируют полевка Microtus obscurus Eversmann и малая мышь (Sylvaemus uralensis Pallas), значительно ниже значение как прокормителей клещей желтогорлой мыши (Sylvaemus flavicollis Melchior), малой белозубки (Crocidura suaveolens Pall.) численность которых не достигает высоких значений. В открытых стациях предгорий прокормителями предимаго могут также быть домовая (Mus musculus Linnaeus) и курганчиковая (Mus hortulanus Nordmann) мыши, общественная полевка (Microtus socialis Pall).
В степных районах, в том числе и на Керческом полуострове, отмечено около 10 видов грызунов, из которых определенную роль в прокормлении клещей играют виды-доминанты - домовая и степная (Sylvaemus arianus Blanford) мыши, общественная полевка, и в отдельные годы - малая белозубка.
Хозяевами взрослых клещей в горно-лесных районах служат домашние (коровы, козы, овцы) и дикие виды копытных (косули, олени), а также хищники (лисицы, собаки) и зайцы-русаки. В степных районах основные прокормители клещей - домашний скот, лисицы, собаки и зайцы.
Для успешного развития той или иной микропопуляции клещей, и особенно для образования очага их массового размножения, необходим ряд условий. Во-первых, благоприятные гидротермические условия в микробиотопах, где происходит развитие яиц, личинок и нимф, как наиболее уязвимых фаз развития клещей в период с мая по август. Во-вторых, наличие в достаточном количестве мелких млекопитающих, как прокормителей преимагинальных фаз развития, а также средних и крупных млекопитающих - как прокормителей имаго, а частично и нимф.
При этом, если мелкие млекопитающие, встречающиеся повсеместно в большем или меньшем количестве, могут обеспечить прокормление большинства предимаго, то с прокормителями имаго дело обстоит много сложнее. Их численность во многих местообитаниях очень ограничена, поэтому нередко, из перечисленных факторов на образование и функционирование локального очага клещей решающее значение имеет наличие достаточного числа именно прокормителей имаго (самок). В последнее десятилетие идет постоянное и многократное увеличение численности коров, коз, появились и многочисленные отары овец, выпас которых часто ведется в местах концентрации иксодовых клещей - на лесных полянах, опушках, в разреженных кустарниковых зарослях. Все это способствует увеличению численности иксодовых клещей и может привести к активизации очагов ряда трансмиссивных инфекций (клещевого энцефалита, иксодовых клещевых боррелиозов, туляремии и других), в функционировании которых важное значение играют клещи D. reticulatus и D. marginatus.
Развитие клещей
Клещи D. reticulatus и D. marginatus, развитие которых протекает по треххозяинному типу, т.е. со сменой трех хозяев, имеют во многом сходные циклы раз-вития (Олсуфьев, 1953, Daniel et al., 1980; Разумова, 1986, Покровская, 1951, 1953; Nosek, 1972; Gilot, Pautou, 1985).Продолжительность их жизненного цикла, как и у других палеарктических видов этого рода, охватывает один год.
Оптимальными условиями развития D. reticulatus следует считать: температуру 18-250С, влажность 60-80% (Резник, 1970). Отклонения в ту или иную сторону могут ускорять или замедлять развитие клещей.
На все развитие от окончания питания самок до появления имаго следующего поколения требуется 90-110 сут (Балашов, 1998). Период откладки и созревания яиц, занимает от 20-30 дней у D. marginatus до 35-45 дней - у D. reticulatus (Резник, 1970).
Выход личинок р. Dermacentor начинается обычно через месяц после откладки яиц и растягивается на 10-15 дней. Активация личинок наиболее интенсивно происходила в течение первой недели после начала вылупления. Если в лабораторных условиях из одной кладки выходило до 1000 личинок, то в природных условиях это число не превышало 200-400 (Фильчагов, Лебедева, 1988). Наибольшая плотность активных особей в течение всего периода наблюдения сохранялась в радиусе 1-1.5 м от места кладки. Максимальное удаление личинок от места выплода (2.5 м) и наибольшая площадь занимаемой ими территории (8,3 м2) были зафиксированы к концу второй недели после начала выхода.
В Крыму личинки появляются около середины июня, встречаются в природе до конца августа. Напитавшиеся личинки линяют на нимф, которые встречаются в природе с конца июня по август.
В июле-августе начинают появляться имаго нового поколения. Большая часть последних приступает к питанию после перезимовки, хотя наблюдается и осенняя волна активности и паразитирования клещей на животных. В этом случае откладка яиц происходит только у перезимовавших особей через 9-12 сут после насыщения, а вылупление личинок происходит через 12-15 сут.
В районах с более суровым климатом, предимагинальные фазы клещей встречаются в природе на протяжении более короткого периода. Так, например, в Каракалпакии личин-ки и нимфы наблюдаются на мелких мышевидных грызунах с мая по октябрь, при максимальной численности их в июне-июле (Сержанов, 1965). Развитие D. marginatus и D. reticulatus в условиях жаркого лета Крыма, как и южных районов России (Балашов, 1998), происходит в более сжатые сроки. К примеру, метаморфоз личинок занимает 3-7 сут, нимф - 22-26 сут, в то время, как в более северных районах метаморфоз напитавшихся личинок занимает 12-22 сут, нимф - 25-60 сут (Daniel et al., 1980).
Голодные личинки живут в природе не более 20-60 сут и нимфы - 35-70 сут (Покровская, 1951, 1953, Daniel et al., 1980, Ламанова, 1966). Поэтому, из-за эфемерности неполовозрелых форм зимовка обоих видов возможна только на стадиях голодных имаго и напитавшихся самок, а голодные или напитавшиеся, но не перелинявшие к зиме нимфы поги-бают. Напротив, голодные имаго отличаются исключительной долговечностью. Во многом длительность жизни зависит от условий существования и в частности зимовки - в открытых и сухих стациях клещи живут меньше, чем в закрытых с повышенной влажностью. Продолжительность жизни взрослых клещей достигает трех летних сезонов с момента вылупления личинок (Адамович, 1961, Разумова, 1966; Балашов, 1998). При этом в пер-вую зимовку выживает до100% клещей, во вторую - около 80, во время третьей зимовки, все гибнут (Покровская 1958, Ламанова 1966). Вследствие этого активизирующаяся весной или осенью популяция представляет смесь из особей не менее трех поколений.
Время выхода зимующих имаго и их активизация весной во многом зависит не только от климатических факторов данного года, но и от конкретных микроклиматических условий мест зимовки. Повсеместно перезимовавшие имаго активизируются ранней весной со сходом снега и наступлением первых теплых дней.
В Крыму, в природных местообитаниях, успешно перезимовавшие клещи D. marginatus встречаются уже в теплые февральские "окна", а в марте отмечается их максимальная активизация (средне - многолетняя численность - 0.33 экз. на флаго/час) (Рис 2).
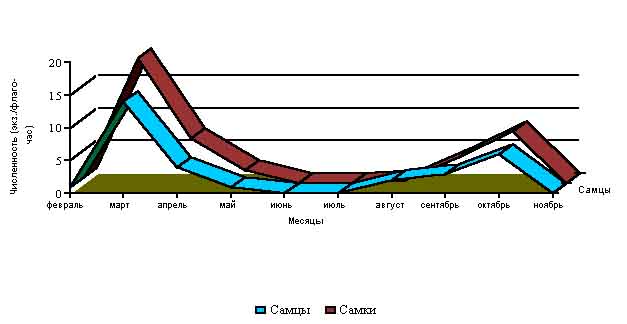 Рис. 2. Динамика численности клещей D.marginatus по месяцам
Рис. 2. Динамика численности клещей D.marginatus по месяцам
(экз/флаго-час.)
В местах с наиболее оптимальными условиями существования встречалось до 50-80 экз. на 1 флаго/час и больше. В апреле численность клещей снижается в три раза по сравнению с мартом, а в июне - еще в три раза по сравнению с маем. Причин такой динамики несколько. Во-первых, самки, нашедшие хозяев и успешно завершившие питание, уходят в подстилку, где начинают откладку яиц, после чего они погибают. Во-вторых, постоянно происходит естественная гибель особей от хищников, неблагоприятных климатических факторов и других причин. В-третьих, постепенно происходит повышение среднесуточных температур, сильно повышается сухость воздуха, что неблагоприятно влияет на активность клещей. В летние месяцы, характеризующиеся жаркой и сухой погодой (июнь - первая половина августа), оставшиеся в живых клещи покидают растительность и укрываются от неблагоприятного воздействия высоких температур, низкой влажности воздуха и сильной инсоляции в подстилке, впадая в летнюю диапаузу. После снижения среднесуточных температур и выпадения дождей, характерных для конца августа, происходит активизация клещей, что выражается в постепенном нарастании их численности вплоть до середины октября. Хотя суммарная численность имаго в популяциях в данный период намного выше таковой чем весной, однако большинство клещей нового поколения не активизируются и не покидают подстилку. С конца октября и в ноябре, когда температура становится отрицательной, клещи снова покидают растительность и укрываются в расщелинах почвы, подстилке и других укромных местах, где и зимуют. Отдельные особи клещей, в связи с нередкими для Крыма теплыми и погожими "окнами", которые случаются в ноябре-декабре, могут активизироваться и встречаться в активном состоянии на остатках травянистых растений и кустарниках.
Динамика численности D. reticulatus аналогична предыдущему виду (см табл. ). Только в связи с тем, что в верхнем поясе гор весной теплая погода устанавливается позже, соответственно и сроки активности клещей смещены на более поздние сроки. Так активизация клещей после зимовки происходит в марте, достигая своего пика в апреле, а с мая снижается, падая почти до нуля. Осенняя динамика активности клещей D. reticulatus примерно соответствует таковой клещей D. marginatus. Осенние волны заклещевления, как пра-вило, бывают менее выраженными, чем весенние. Отличительной особенностью клещей D. marginatus является их способность зимо-вать на хозяине, в то время как зимующих D. reticulatus на животных не обнаружено (Резник, 1970). Сборы, сделанные в холодное время года, мо-гут быть довольно обильными и содержать напившихся самок (зимовать остаются самки, присо-савшиеся поздно осенью) (Галузо, 1948).
Близкие данные по фенологии клещей отмечены и в других районах ареала (Адамович, 1961, Галузо, 1948, Разумова 1962, Ганиев, 1956). Однако иногда отмечаемое исследователями превышение осеннего пика численности над весенним (Сапегина, 1965, Богданов и др., 1964), в Крыму не наблюдалось. Длитель-ность обеих волн сезонной активности более или менее одинакова и продол-жается около двух месяцев (Логиновский, l966).
Поведенческая и репродуктивная диапаузы регу-лируются длиной светового дня. Для D. marginatus характерна короткодневная фотопериодическая реакция с критическим фотопериодом в 13 ч (Бе-лозеров, 1976). Экологическое значение обеих форм диапаузы однозначно и на-правлено на предотвращение откладки яиц в конце лета и осенью и гибели не достигнувших имагинальной фазы особей зимой. Самки, напитавшиеся с середины лета, и осенью находятся в состоянии репро-дуктивной диапаузы и приступают к яйцекладке только после зимовки вместе с особями весеннего насыщения В июне-июле клещи становятся неактивными и прекращают нападать на хозяев, хотя и могут оставаться на растительности. В сезонной регуляции главная роль принадлежит имагинальной диапаузе (Балашов, 1998).
Суточная активность
Половозрелые особи клещей р. Dermacentor суточных миграций практически не совершают, оставаясь на выбранных растениях постоянно. Это подтверждают специальные наблюдения, проведенные на Ай-Петринской яйле во второй декаде сентября. Клещи оставались на растительности в позе пассивного ожидания в течение всех суток, несмотря на резкие перепады температуры. Они одинаково активно цеплялись на флаг днем, когда температура воздуха достигла +150С; поздно вечером, после наступления сумерек при 5-70С тепла и рано утром, при падении температуры воздуха до 30С ниже нуля.
Сезонная регуляция одногодичного цикла обеспечивается существованием летней неативности у голодных и диапаузы у напитавшихся самок. Впадение клещей этих видов в состояние летней неактивности и возникновение несколько позднее потенциальной способности к диапаузе у напитавшихся самок служит надежной биологической адаптацией, предотвращающей появление в конце лета и осенью не способных к зимовке неполовозрелых фаз развития (Фильчагов, Лебедева, 1988).
Как показали наблюдения этих же авторов, прокормители собирают 10-20 активизировавшихся в агрегациях личинок. Разброс напитавшихся особей составляет около 1 га. Единичные заносы отмечены на расстояние до 300 м. Однако в отдельных случаях возможен занос птицами или более подвижными животными (например зайцем-русаком) и на более дальние расстояния.
Индивидуальные различия массы взрослых клещей (как и их размеры), зависят в основном от количества крови поглощенной на предимагинальных стадиях (в большей мере на нимфальной, в меньшей - на личиночной) и длительности вынужденного голодания в ожидании прокормителя. В третьей декаде марта 1988 года, в трех различных биотопах нами были собраны клещи, вышедшие с мест зимовки. Сравнение полученных данных по индивидуальному взвешиванию живых клещей показало, что масса отдельных особей самцов колебалась от nmin=3,5 мг до nmax =18 мг, при средней массе самцов в трех выборках: 8,4+0,4; 9,4+0,9; 9,4+1,4 мг. Масса самок изменялась от nmin=3,0 мг до nmax =14,0 мг, при средней массе самок в трех выборках: 8,1+0,2; 7,5+0,3; 8,4+0,4 мг. Во всех трех локальных популяциях самцы достоверно крупнее самок и эта разница (по средним показателям) колеблется от 4,1% до 19,9%, что связано с наличием у самцов сильно хитинизированного покрова. Длина тела имаго в голодном состоянии колебалась в пределах 2-8,5 мм.
Особенности поведения.
Биологическая особенность клещей р. Dermacentor - их способность "притворяться мертвыми" после какого-либо беспокойства (прикосновения к клещу и т.п.). Эта способность свойственна, далеко не всем видам клещей и, например, она отсут-ствует у клещей рода Ixodes, но наблюдается у некоторых видов р. Hyalomma. По-видимому, эта особенность поведения является биологически целесообразной у тех животных, которые, упав с растений, попадают не в густую растительность или почвенную подстилку, а на открытое место.
Виды р. Dermacentor в позе "ожидания" хозяина располагаются на стеблях растений всегда передним концом тела вниз, в отличие от клещей р. Ixodes, сидящих на растениях передним концом вверх. В случае обнаружения приближающегося позвоночного, они сначала широко расставляют в стороны последнюю пару лапок, а перед непосредственным контактом - первую, а иногда и вторую, при этом удерживаясь на стебельке только третьей парой лапок.
Поведение личинок резко отличается от такового взрос-лых клещей. В то время как имаго сидят на расте-ниях в позе "ожидания", личинки (D. marginatus) никогда не поднимаются на стебельки трав (Галузо, 1948). Они довольно активно, в зависимости от температуры окру-жающей среды, ползают по поверхности земли, по сухим расти-тельным остаткам, заползают в трещины почвы и в норы грызунов. По-видимому, такое поведение является причинной того, что предимаго клещей этого рода не попадаются при собирании их с растений при помощи флага или сачка. Эта особенность поведения личинок позволяет им в усло-виях сухого климата скрываться от иссушающего действия солнца, а такой способ "охоты" обеспечивает им более быстрое обнаружение будущего хозяина и прикрепление к нему. Возможно, что такие особенности поведения и могли привести к выработке и к закреплению впоследствии у предимагинальных фаз клещей их связи именно с мелкими млекопитаю-щими, а так как возможностей встречи с птицами у личинок очень мало, поэтому на пернатых молодые фазы клещей этого рода почти не паразитируют.
Выводы
1. В фауне Крыма иксодовые клещи рода Dermacentor представлены двумя видами: D. marginatus, ареал которого занимает практически весь полуостров, и D. reticulatus, образующий в Крыму географически изолированную от основного ареала популяцию, ограниченную горно-лесной зоной.
2. Оба вида клещей характеризуются определенной гигрофильностью, предпочитая лесные экотонные сообщества (лесные поляны, опушки леса, лесосеки и т.п.) горного Крыма с развитым травянистым покровом, где образуют очаги массового размножения.
3. Распределение клещей в пределах ареала носит ярко выраженный агрегированный характер. В горно-лесной зоне отмечены определенные вертикальные закономерности в распределении клещей на высотах от 0 до 1500 м над уровнем моря.
4. Прокормителями предимаго клещей служат в основном мелкие млекопитающие (общественная и обыкновенная полевки; малая, степная, желтогорлая, домовая и курганчиковые мыши, малая белозубка и др.). Прокормители имаго - домашний скот, дикие копытные (олени, косули), зайцы, лисицы, собаки и др. животные.
5. Развитие клещей протекает по треххозяинному типу и завершается в течение года. Имаго отличаются исключительной долговечностью (живут до трех лет), личинки и нимфы существуют в природе короткий период (до 2-2,5 месяцев). Зимуют только имаго.
6. Отмечено два пика сезонной активности: весенний и осенний. Суточная активность у клещей не выражена.
1. Адамович В. Л. Ландшафтно-географическое размещение иксодовых клещей в Волынском Полесье. // Зоол. журн., 1961, т. 10 , вып.5.- С. 676-685.
2. Балашов Ю. С. Иксодовые клещи-паразиты и переносчики инфекций // Санкт-Петербург.: Наука- 1998.- С. 287.
3. Белозеров В. Н. Жизненные циклы и сезонные адаптации у иксодовых клещей (Acarina, Ixodoidea) // Докл. на XXVIII ежегод. чтении пам. Н. А. Холодковского. Л., 1976. - С. 53-101.
4. Богданов И. И., Алифанов В. И., Федоров .В. Г. // Некоторые факторы изменения численности иксодовых клещей в Западной Сибири. // В сб.: Совр. проблемы изучения динамики численности популяций животных: 1964.- С. 46-47.
5. Вшивков Ф. Н. Иксодовые клещи диких позвоночных животных Крыма // Девятое совещание по паразитол. проблемам. Тез. докл.- М.-Л.:Изд-во АН СССР.- 1957.- С. 41-42
6. Вшивков Ф. Н. К фауне и экологии иксодовых клещей диких позвоночных животных Крыма // Изв. Крьмск. пед. ин-та. -1958.- т. XXXI.- Симферополь -С. 47-61 .
7. Галузо И. Г. Физические факторы местообитания иксодовых клещей. Изв. // АН КазССР, сер. паразитол., 1948, 43 (5). - С. 3-12.
8. Ганиев И. М. К биологии клеща Dermacentor marginatus Sulz. в Дагестане. // Тр. ин-та животноводства Даг. фил. АН СССР, 1956, 4. - С. 248-254.
9. Геоботанiчне районування Украiнськоi РСР // К.: Наукова думка.- 1977.- С. 302
10. Гутова В. П., Сиротюк Н. П. Энтомологическая ситуация в очагах клещевого энцефалита в Крыму // 12-я Всесоюз. конф. по природной очаговости болезней. Тез. докл. (10-12 окт. 1989 г, Новосибирск). М., 1989. С. 53-54.
11. Емчук Е.М. Фауна Украины. Иксодовые клещи.// Т.25, вып.1. Изд-во АН УССР, К., 1960.- С. 163.
12. Емчук Е.М. Матерiали до фауни i екологii iксодових клiщiв Кримськоi областi. // Працi iнституту зоологii АН УРСР, т. XIV.- К.- 1957.- С. 3-17.
13. Кочкин М.А. Почвы, леса и климат Горного Крыма и пути их рационального использования // Науч. тр. Никитского бот. Сада. М.: Колос.- 1967.-Т.38.- С.368.
14. Ламанова А. И. Схемы размножения Dermacentor marginatus // Уч. зап. Карагандинского мед. ин-та, 1966, 3.- С.
15. Логиновский Г. Е. К экологии клеща Dermacentor pictus Herm. в Курганской области. // Фауна и экология членистоногих Сибири. Новосибирск.1966.- С. 28-29.
16. Маркешин С. Я., Караванов А. С., Ковин В. В., Захарова Т. Ф., Евстратов Ю. В. Бычкова М. В,, Евстафьев И. Л. Очаги клещевого энцефалита в Крыму // 12-я Всесоюз. конф. по природной очаговости болезней. Тез. докл. (10-12 окт. 1989 г., Новосибирск). М., 1989.- С. 100.
17. Маркешин С. Я., Смиронова С.Я., Евстафьев И. Л. Оценка состояния природных очагов Крымской-конго геморрагической лихорадки в Крыму // Журн. микробиол. - 1992.- ⌡4.- С.28-31.
18. Маркешин С.Я. Изучение очагов клещевого энцефалита, геморрагической лихорадки с почечным синдромом и крымской-конго геморрагической лихорадки в Крыму. // Диссертация канд. мед наук., М.,-1994,- С. 160.
19. Мельникова Т. Г. Иксодовые клещи диких и домашних животных Крымского заповедника. // Зоол. журн.., т. 32, в.3. 1963.- С. 452-458.
20. Мельникова Т. Г. О развитии и распространении клеща Dermacentor marginatus Sulz в условиях горно-лесного Крыма. // Зоол. журн., т. 10, вып. 6, 1961.- С. 713-717.
21. Мельникова Т. Г. Иксодовые клещи Крымского заповедника. // Труды Крымского филиала АН УССР, т. IX, Зоология, вып. 3.- Симферополь 1955.- С. 61-68.
22. Олсуфьев Н. Г. О происхождении очагов клеща Dermacentor pictus Herm. в южной части Московской области. // Тез. докл. Второго совещания по паразитол. проблемам: 1940,- С. 26-28.
23. Олсуфьев Н. Г. К экологии лугового клеща Dermacentor pictus Herm., о происхождении его очагов и путях их ликвидации в средней полосе Европейской части РСФСР // Вопросы краевой, обшей, экспериментальной паразитологии и медицинской зоологии. М. 1953. Т. 8.- С. 49-98.
24. Покровская В. И. К экологии клеща Dermacentor marginatus в Воронежской области // Зоол. журн. 1953. Т. 32. вып. 3.- С. 435-440.
25. Покровская В. И. К экологии личинок и нимф клеша Dermacentor marginatus в условиях Воронежской области // Зоол. журн. 1951. Т. 30, вып. 3.- С. 224-228.
26. Покровская Е. И. К вопросу о летней пассивности половозрелых клещей Dermacentor marginatus Sulz. и о длительности их голодания в условиях Юго-Востока Черноземного центра.// Мед. пар. и пар. бол., 1958,4.- С. 487-488.
27. Померанцев Б.И. Фауна СССР. Иксодовые клещи (Ixodidae) // Т.1У, вып.2. Изд-во АН СССР. М.-Л., 1950.- С 224.
28. Померанцев Б. И. Матикашвили Н. В. Эколого-фаунистический очерк клещей Ixodidae (Acarina) Закавказья. // Пар. сб., 1940, 7.- С. 100-133.
29. Разумова И. В. Определение физиологического возраста и возрастной состав природной популяции Dermacentor pictus Herm. // Мед пар и пар бол., 1962. 1.- С. 55-61.
30. Разумова И. В. Длительность жизни популяции голодных половозрелых клешей Dermacentor pictus Herm. // Мед. паразитол. 1966. 3.- С. 293-298.
31. Разумова И. В. Физиологический возраст иксодовых клещей (понятие, методы определения, популяционно-экологические вопросы, перспективы прикладного использования) // Автореф. дис. канд. биол. наук. М., 1986.- С. 23.
32. Резник П. А. Особенности ареалов и пути формирования фауны иксодовых клещей Советского Союза. // Фауна Ставрополья. Ставрополь 1970.- С. 3-187
33. Сапегина В. Ф. Экология иксодовых клещей лесостепного очага клещевого энцефалита северо-восточного Алтая. // В сб.: Природа очагов клещевого энцефалита на Алтае. Новосибирск 1965.- С. 38-42.
34. Сержанов О. С. Иксодовые клещи (Ixodoidea, Parasitiformes) Каракалпакии и их эпидемиологическое и эпизоотологическое значение. // Автореферат дисс. на соиск. уч. степ. канд. биол. н. Нукус. 1965.- С. 1-27.
35. Фасулати К.К. Полевое изучение наземных беспозвоночных // Изд-во "Высшая школа".-М.-1971.- С. 424.
36. Федюшин А. В. Клещ Dermacentor pictus Herm., 1804 - переносчик очаговых заболеваний человека и домашних животных в условиях Омской областа // Тр. Омского с.-х. ин-та, 1960, 40.- С. 157-180.
37. Фильчагов А.В., Лебедева Н.Н., К изучению экологии голодных личинок Dermacentor reticulatus и их связей с прокормителями в естественных условиях // Паразитология, 22, 5.- 1988.- С. 366-371.
38. Daniel M., Szymanski S., Cerny V., Dusbabek F., Honwkova E., Olejnicek J. A comparison of developmental dynamics of Dermacentor reticulatus of different geographic origins and their affection by different microclimate // Folia Parasitol. 1980. Vol. 27, N 1.- P. 63-69.
39. Gilot В., Pautou C. Repartition et ecologie de Dermacentor marginatus Sulzer, 1776 (Ixodoidea) dans les Alpes francaises et leur avant-pays // Acarologia. 1985. Vol. 24. N 3.- P. 261-273.
40. Nosek J. The ecology and public health importance of Dermacentor marginatus and D. reticulatus ticks in Central Europe // Folia Parasitol. 1972. Vol. 19, N 1.- P. 93-102.