Пролетные пути и эволюция птиц
В.Н. Грищенко
г. Канев, Каневский заповедник
Дискуссия о том, как летят птицы - широким фронтом или по узким пролетным путям, точилась многие десятилетия (подробнее об этом см.: Кумари, 1957; Грищенко, 1994а). Сейчас большинство ученых склоняется к точке зрения, которую можно назвать "синтетической": миграция идет широким фронтом, но внутри него образуются участки с повышенной концентрацией мигрантов - пролетные пути, которые могут соединяться и разветвляться. Х. Гейр фон Швеппенбург (Geyr von Schweppenburg, 1931) предложил для устранения разнобоя в трактовке использовать вместо последнего термина название "узкий фронт". Оно прижилось и часто употребляется, но важно не забывать, что узкий фронт не адекватен пролетному пути в современном его понимании. Различие состоит в том, что за границами узкого фронта мигранты практически не встречаются. Пролетный путь же лишь "сгущение" внутри широкого фронта миграции. Птицы летят и за его пределами, но в меньших количествах. Таким образом, всякий узкий фронт миграции является пролетным путем, но не всякий пролетный путь - это узкий фронт.
1
Американский орнитолог Ф. Белроуз (Bellrose, 1968) на основании многолетних комплексных исследований миграции водоплавающих птиц в США (кольцевание, радарные и визуальные наблюдения, прослеживание с помощью самолета) предложил выделить пролетные пути различных уровней. Пролетный путь высшего порядка, который прослеживается в масштабах континента, - flyway. Адекватно перевести его на русский язык не так просто, потому что сам термин flyway (в широком смысле - пролетный путь) нередко употребляется в различных значениях, к тому же по разному переводится. Например, в книге К. Карри-Линдала (1984) он фигурирует как миграционный поток, хотя последний термин часто употребляется у нас в других значениях. Такой пролетный путь можно назвать континентальным, поскольку он прослеживается в пределах 1-2 континентов, непосредственно связывая места гнездования с зимовками. Миграция птиц в Северной Америке хорошо изучена, и там выделено 4 таких континентальных пролетных пути: Атлантический, Миссисипский, Центральный и Тихоокеанский (Lincoln, 1935). Дальнейшими исследованиями было установлено, что в их пределах выделяются пути более низкого порядка. Ф. Белроуз (1968) называет их миграционными коридорами (migration или flight corridors). Эти коридоры ведут от конкретных мест гнездования. Они сливаются, образуя более мощные миграционные потоки, связывают между собой различные континентальные пролетные пути. Последние представляют собой причудливую сеть из миграционных коридоров. Различные виды птиц могут использовать разные коридоры. Те в свою очередь состоят из сети пролетных путей более низкого уровня (migration routes). Ширина их может достигать всего несколько километров. В плане терминологии нам кажется вполне приемлемым предложение Э. Шюца (Schüz, 1968) называть описанные образования пролетными путями I, II и III порядков. Можно назвать также два последних уровня соответственно региональными и местными пролетными путями.
Такие пролетные пути трех уровней хорошо узнаваемы и для миграции европейских птиц. Проследим это на примере белого аиста (Ciconia ciconia). Континентальный пролетный путь или flyway - это путь миграции восточной или западной популяций в целом (рис. 1). Пролетные пути, выделяемые в пределах отдельных регионов, например, Украины, - не что иное, как миграционные коридоры Ф. Белроуза. Их сеть также образует континентальный пролетный путь (для одного вида его можно было бы также называть популяционным, поскольку речь идет о миграции в целом конкретной популяции). То, что они распадаются на более мелкие образования, было показано нами на примере скворца (Sturnus vulgaris) (Грищенко, 1992).
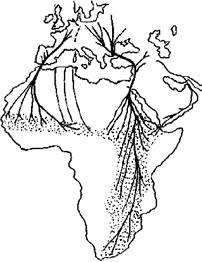
Рис. 1. Миграции белого аиста (по: Schulz, 1988 с дополнениями).
2
Мы видим, что сеть пролетных путей различных порядков образует внутри миграционного ареала своеобразную упорядоченную структуру. Как связано ее появление с эволюцией птиц вообще, возникновением и развитием миграций в частности?
Известно, что динамическая система, имеющая упорядоченную внутреннюю структуру, тратит в целом меньше энергии. Мигрирующую популяцию птиц также можно рассматривать как такую систему. Естественно, этой популяции выгодно в эволюционном плане тратить минимум энергии на достижение зимовок и мест гнездования. Насколько это важно, хорошо иллюстрирует такой пример.
Исследованиями Й.Х. Даллинги и С. Шёнмакерса (Dallinga, Schoenmakers, 1989) показано, что динамика численности и успешности размножения белого аиста в Западной Европе тесно коррелирует с количеством осадков на местах зимовки в Африке, которые определяют состояние кормовой базы. После зим с обильными дождями аисты прилетают на места гнездования раньше, успешность размножения в такие годы выше. Другими словами, успешнее размножаются те птицы, которые имели лучшие энергетические ресурсы. Значительно влияет на это также расход энергии на путях пролета.
Таким образом, та популяция данного вида, которая вернется в места гнездования с наименьшим расходом энергии, будет иметь большую успешность размножения (конечно на это влияют и условия в местах гнездования, но состояние птиц к моменту прилета является очень важным), т. е. такая "экономная" миграция будет закрепляться в ходе эволюции. По нашему мнению, возникновение и существование системы пролетных путей имеет под собой именно энергетическую основу, которая приводит в действие эволюционный механизм.
Пролетный путь - это участок миграционного ареала вида, где миграция может проходить с минимальными затратами энергии, т. е. оптимальный путь в данных условиях. Всякое отклонение от этого оптимального пути (естественно, на его расположение влияет очень много факторов, в том числе изменчивых, и местонахождение такого оптимума тоже может смещаться) ведет к увеличению затрат энергии, поэтому естественный отбор будет оказывать стабилизирующее воздействие на упорядоченность структуры миграционного ареала.
В чем же может проявляться оптимальность миграции по упорядоченной системе пролетных путей? На их расположение влияет ряд факторов.
1) Возможность миграции вообще данного вида в данном месте. Хорошо известно, что узкоспециализированные наземные парители - белый аист, крупные хищники - избегают пересекать большие морские пространства и огибают их над сушей. В тех случаях, когда миграция над экологическим барьером невозможна или ведет к чрезмерным затратам энергии, образуется узкий фронт пролета с огромной концентрацией мигрантов на ключевых участках (например, Босфор или Гибралтар).
2) Оптимальность условий миграции в данном месте. Определенные участки оказываются привлекательными для птиц в силу аэродинамических, защитных и др. факторов. Парители концентрируются вдоль горных цепей и гряд холмов, водоплавающие птицы часто следуют вдоль водоемов, мелкие дендрофильные птицы могут избегать перелетать большие открытые пространства. Так осенью образуется поток мигрантов, огибающих Киевское водохранилище (Полуда, 1983). Большое значения для формирования пролетных путей имеют ландшафтные ориентиры (Михеев, 1992). Во многих из подобных случаев мы имеем дело с эффектом направляющих линий. Влияют и погодные условия в данной местности. Весной многие птицы, например, белый аист, более охотно огибают Карпаты, образуя два обтекающих их миграционных потока, осенью же пролетный путь пересекает горную цепь (Грищенко, Серебряков, 1992). Понятно, что весной условия полета над горами, где снег лежит весь период миграции, гораздо хуже, чем осенью, когда устойчивый снежный покров еще отсутствует.
Во всех этих случаях давление факторов среды не так сильно, как в первом из рассмотренных вариантов, и птицы могут мигрировать в различных местах. Но все же достаточно четко прослеживается статистическая закономерность: большинство их мигрирует там, где это энергетически выгоднее. Предпочтут ли мигранты огибать или пересекать экологический барьер, определяется многими факторами. Но прежде всего играет роль соотношение энергетических затрат на более прямой, но трудный, и более длинный, но легкий из путей. Так, стае зябликов (Fringilla coelebs) ничего не стоит облететь Киевское водохранилище, оставаясь в привычных условиях, но Черное море им уже гораздо проще пересечь напрямую. Тут следует учитывать и индивидуальное состояние мигрантов. Птицы в высокоразвитом миграционном состоянии с большими жировыми резервами могут пересекать даже весьма протяженные экологические барьеры (например, Средиземное море + Сахара или Мексиканский залив, миграция сухопутных видов над Атлантическим океаном). Но для птиц с недостаточными жировыми запасами это уже может быть невозможным. Здесь проявляются две различные стратегии миграции - преодоление больших расстояний единым броском и "короткими перебежками". Количественное соотношение между видимой и высотной миграциями в данном случае не играет роли. Важно то, что в целом для популяции выгодно существование обходных путей.
3) Минимальность расстояния. Часто миграционные потоки проходят кратчайшим путем от одних ключевых участков к другим. Энергетическая выгода здесь очевидна.
4) Оптимальность кормовых условий местности. Это очень важно прежде всего для птиц, использующих стратегию "коротких перебежек", но и для мигрантов, пролетающих большие расстояния, тоже необходим длительный отдых и кормежка. Иногда этот фактор является решающим в определении положения пролетных путей. Хороший пример тому - миграция белого аиста на Ближнем Востоке (см. ниже). Птицы летят узкой полосой только там, где возможна кормежка (после пересечения больших участков горной местности на Балканах и в Малой Азии это особенно важно).
В тех случаях, когда имеет место длительный беспосадочный перелет (например, над морем), птицы могут лететь более или менее равномерно, хотя здесь также возможно образование сгущений, т. е. пролетных путей, связанных с расположением пролетных путей на суше (фактор минимального расстояния) или особенностями атмосферной циркуляции. Устойчивый попутный ветер приводит к значительно меньшим затратам энергии. Может играть роль также расположение ориентиров на противоположном берегу (опять-таки это снижает потери энергии).
Теперь перейдем к рассмотрению образования пролетных путей на конкретных примерах.
3
В Европе существует две популяции белого аиста, которые имеют различные пути пролета и места зимовки (рис. 1). Само их возникновение связано с необходимостью облета Средиземного моря. Линия раздела проходит через Голландию, Гарц, Баварию, Альпы. Западная популяция мигрирует осенью на юго-запад через Францию, Испанию, Гибралтар. Зимует она в саваннах Западной Африки. Восточная популяция мигрирует на юго-восток и юг. Небольшие пролетные пути сливаются на Балканах в большую магистраль, которая ведет к Босфору. После него птицы направляются на юго-восток и пересекают Малую Азию напрямую "по диагонали". Возле залива Искандера они выходят к Средиземноморскому побережью, после чего резко изменяют направление миграции и сворачивают на юг (Creutz, 1988; Schulz, 1988).
Пролет аистов в районе залива Искандера хорошо иллюстрирует воздействие вышеупомянутых факторов на образование и расположение пролетных путей. Это одна из ключевых точек на миграционных путях восточной популяции, и там проводились специальные исследования в 1963-1965 гг. (Heckenroth, 1968). Часть аистов облетает залив (30-40 км шириной), но большинство пересекает его в верхней части (рис. 2). Определяющим фактором здесь часто является сила и направление ветра на малых высотах. На восточном берегу залива находится горный хребет Аманус с высотами до 2220 м. Залив аисты перелетают низко над морем - всего 1-1,5 м, для преодоления гор им нужно набрать высоту, поэтому энергетически наиболее выгодно сделать это там, где высота гор минимальна. И действительно, подавляющее большинство их летит в направлении перевала Белен (700 м) у города Искендерун. Образуется полоса шириной порядка 10 км, по которой летят десятки тысяч птиц. При этом большинство их направляется не на сам перевал, а набирает высоту в восходящих потоках над склонами севернее его. За Беленом находится оз. Амик и богатые кормом болотистые местности. После пересечения хребта Аманус аисты попадают в зону устойчивых северных ветров. В этом же районе концентрируются в большом количестве и мигрирующие хищные птицы, которые подобно аистам летят из Европы через Босфор и Малую Азию. Здесь к тому же к ним присоединяется большой поток птиц, пролетающих между Черным и Каспийским морями (Cameron et al., 1967).
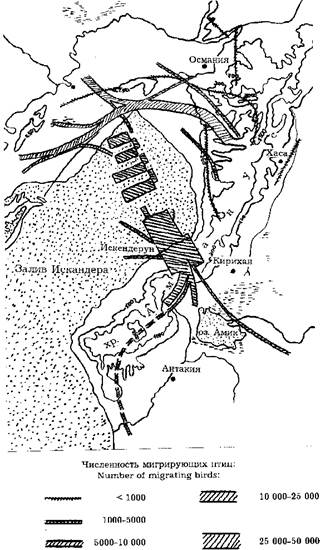
Рис. 2. Осенняя миграция белого аиста в районе залива Искендерун (по: Heckenroth, 1968).
Известно, что вдоль восточного побережья Черного моря идет очень интенсивный пролет хищных птиц. Почему они концентрируются именно здесь, а не в другом месте? Многие хищники также вынуждены огибать морские пространства. Один взгляд на карту показывает, что это кратчайший путь для них из Европейской части России к пролетным путям на Ближнем Востоке. Там же птицы скапливаются в свою очередь потому, что это прямой путь к африканским зимовкам. По восточному побережью Черного моря идет и пролетный путь белого аиста, хотя и слабо выраженный (Лебедева, 1979). Он ведет из территорий с низкой его численностью. Аисты же из Закавказья и Средней Азии мигрируют уже в другом направлении. Поэтому с севера на Ближний Восток летят из парителей главным образом хищники.
Дальше на юг аисты летят узким потоком по долине р. Иордан, через Мертвое море и выходят в верховья залива Акаба. Здесь птицы сворачивают на юго-запад. Большинство их пересекает Суэцкий залив в районе Эль-Тор, часть отклоняется дальше к югу. Достигнув африканского побережья, аисты выходят в долину Нила возле Кены (Koch et al., 1966; Safriel, 1968; Schüz, 1971). На Ближнем Востоке они придерживаются осенью древнего разлома, который проходит под Иорданом и Акабой. Здесь в долине наиболее благоприятные кормовые условия (Koch et al., 1966). Дальше на восток в Сирийской пустыне аисты встречаются на пролете крайне редко (Cameron, Cornwallis, 1966; Safriel, 1968). Лишь в саваннах Судана миграционный поток их начинает расширяться, в Восточной Африке образуются многочисленные ответвления, которые ведут к различным местам зимовки.
Часть аистов из Средней Азии мигрирует напрямую через Аравийский п-в. Путь через пустыню не легкий, но все же он энергетически выгоднее, чем облет через Синай.
Таким образом, мигрирующие белые аисты собираются по "тропинкам" и коридорам в общий магистральный пролетный путь, который идет от Балкан до долины Нила и затем снова распадается на коридоры и "тропинки".
На приведенном примере хорошо видно действие 4 вышеназванных факторов, способствующих образованию пролетных путей. Аисты не могут парить над морем, поэтому образуется мощные обходные потоки вокруг Черного и Средиземного морей. Они могут парить над пустыней на Ближнем Востоке, но кормовые ресурсы там сконцентрированы в очень узкой полосе древнего разлома, и миграция идет снова плотным потоком именно там. Горы Аманус большинство птиц пересекает в энергетически оптимальном месте, за которым к тому же расположены богатые кормом биотопы. Дальнейшее направление миграции определяется расположением горного хребта и зоной устойчивых северных ветров, а затем цепью кормовых биотопов.
Наибольшая концентрация миграции аистов и хищников происходит на восточном побережье Средиземного моря. Это вызывается экстремальным окружением - море с одной стороны и пустыня с другой. Т. е., как видим, упорядочивание структуры миграционного ареала есть реакция птиц на неблагоприятные условия окружающей среды. И это упорядочивание будет тем четче выражено, чем сильнее такое воздействие. Другой вариант - преодоление подобной местности одним броском. Но, подчеркнем еще раз, поскольку не все мигранты могут использовать эту стратегию, для популяции в целом образование таких пролетных путей выгодно в эволюционном плане. При ослаблении неблагоприятного воздействия они снова расширяются. Так, во время осенней миграции белые аисты практически не встречаются на побережье Израиля и Ливана. Весной же пролет по побережью хорошо выражен. Связано это с тем, что в это время здесь стоит влажная погода, и птицы находят достаточное количество пищи (Schüz, 1971). Более широким потоком аисты пересекают весной и Суэцкий залив (Safriel, 1968). Осенью полоса миграции хищников на Ближнем Востоке шире, чем аистов, им проще прокормиться в засушливой местности (Safriel, 1968).
Другой пример - птица с активным полетом, серый журавль (Grus grus). Ему не нужно облетать море, как аисту, пересекают журавли и горы, например, Кавказ. Но часть птиц из района Балтийского моря мигрирует на юго-запад через Германию, Францию и Испанию (Norrevang, 1959) (рис. 3). Пролет идет полосой шириной от 300 до 400 км (Libbert, 1936; Schuz, 1971; Sanden, 1983). Пиренейский п-в журавли пересекают с северо-востока на юго-запад, часть зимует на юго-западе Испании и на юге Португалии, остальные довольно узким потоком направляются через Гибралтар в Африку (Bernis, 1960). Очевидно, в данной ситуации миграция в обход энергетически выгоднее, чем перелет напрямую через Альпы и Средиземное море, за которыми к тому же лежит Сахара. Как видно из рис. 3, Альпы журавли облетают с двух сторон, направляясь на Пиренеи и Балканы. Образование узкого фронта пролета в Западной Европе также не сложно объяснить: миграция идет кратчайшим путем к Гибралтару, и большое отклонение от пролетного пути будет приводить только к лишним энергетическим затратам. Дополнительный расход энергии "окупается" лишь в том случае, если он связан с полетом к местам отдыха и кормежки, но их также выгоднее использовать как можно ближе к основной пролетной магистрали.
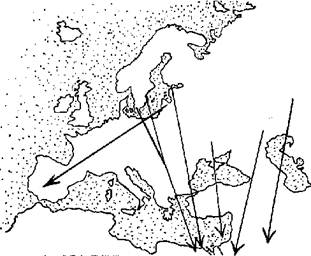
Рис. 3. Пролетные пути европейской популяции серого журавля (по: Norrevang, 1959).
Искушенный читатель скажет, что вышеприведенные примеры описывают лишь переходы широкого фронта миграции в узкий, но не пролетные пути внутри широкого фронта. Да это так. Здесь мы имеем дело с крайней формой образования пролетных путей. У белого аиста узкий фронт пролета образуется в силу невозможности миграции широким фронтом, если можно так выразиться - "запрета среды" на миграцию за пределами пролетных путей. Конечно, бывают исключения: иногда наблюдаются мигрирующие стаи над Кипром (Schüz, 1980) или в пустынях Сирии (Cameron, Cornwallis, 1966), но это именно исключения, связанные к тому же часто с нарушениями ориентации, заносом ветром и т. п. Миграция белого аиста была использована нами как очень удобный пример для иллюстрации закономерностей образования пролетных путей.
Что же будет в том случае, если давление среды на ход миграции оказывается менее жестким? Действие факторов, способствующих возникновению пролетных путей, будет в той или иной степени все равно проявляться. Давление факторов среды приводит к образованию вероятностного распределения признака с более или менее выраженным оптимумом. В нашем случае таким результативным признаком является плотность потока мигрантов. Пики на кривой ее распределения и будут соответствовать пролетным путям, как участкам с повышенной концентрацией птиц. Серый журавль - хороший прример перехода от жестко обусловленного расположения пролетных путей до миграции широким фронтом. В природе существуют как крайняя форма образования пролетных путей - миграция узким фронтом, - так и многочисленные промежуточные варианты. Вполне вероятно, естественно, существование и другой крайности - перелет более или менее равномерным широким фронтом без образования выраженных пролетных путей.
4
Рассмотрим в заключение образование пролетных путей на территории одного региона на примере Украины. Схемы их размещения представлены на рис. 4 и 6. Карта для осенней миграции составлена по нашим данным (Грищенко, 1994б), для весенней - по расположению миграционных потоков на фенологических картах прилета ряда видов птиц (Грищенко, Серебряков, 1988, 1990, 1992, 1993а, 1993б; Серебряков, 1989; Serebryakov et al., 1991). Существование и размещение всех их может быть объяснено действием описанных выше факторов.
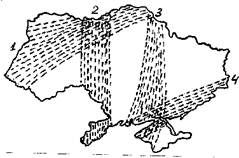
Пролетные пути: Flyways:
1 - Северо-Западный North-West
2 - Бессарабский Bessarabian
3 - Крымский Crimean
4 - Приморский Seaside
Рис. 4. Схема основных пролетных путей весенней миграции птиц на Украине.
Весной на территории Украины образуются 4 основных пролетных пути (рис. 4). Это региональные пролетные пути, миграционные коридоры по терминологии Ф. Белроуза (1968) или пролетные пути второго порядка по терминологии Э. Шюца (1968). У отдельных видов их расположение может несколько отличаться, могут образовываться ответвления, но общие закономерности остаются. Ход миграции в западной половине Украины в значительной степени определяется облетом птицами Карпат. Образуются два больших потока, огибающие горы с запада и востока. О них писал еще А.А. Браунер (1916). Часть мигрантов, естественно, летит напрямую и пересекает хребет, но для популяций перелетных птиц выгодно в эволюционном плане иметь обходные пути через богатые кормом Паннонскую низменность и низовья Дуная. Это значительно снижает смертность во время миграции. Прохождение пролетного пути через Крым (впрочем не у всех видов он пролегает через сам полуостров) помогает птицам сократить продолжительность миграции над открытым морем. Но более важным, возможно, является другой фактор. На рис. 5 приводится схема основных траекторий переноса воздушных масс над территорией Украины. Траектории, проходящие через северные области Украины и низовья Днепра очень хорошо совпадают с Северо-западным и Крымским пролетными путями.

Рис. 5. Схема траекторий переноса воздушных масс в районе Украины (по: Борисов, 1975).
Объяснимы также и отклонения в расположении пролетных путей у отдельных видов. Так, у черного коршуна (Milvus migrans) весной миграционный поток через центральные области Украины смещается к востоку от Крыма, дальше, чем у других птиц (Грищенко, Серебряков, 1990). Связано это с тем, что коршун, как и многие другие хищники, предпочитает мигрировать ближе к побережью Черного моря. Подобно этому, у белого аиста осенью отсутствует Крымский пролетный путь. Вместо него образуется другой, идущий на юго-запад через центральные области Украины к западному побережью Черного моря (Грищенко, Серебряков, 1992).
Расположение осенних пролетных путей тоже определяется действием внешних факторов (рис. 6). Западный - это кратчайший путь из Белоруссии и Прибалтики к "воротам" между Карпатами и Черным морем. Он сливается с двумя другими пролетными путями, образуя мощный поток вдоль западного побережья Черного моря. Подобно этому существование Восточного пролетного пути связано с концентрацией птиц вдоль восточного побережья Черного моря. По нему летят мигранты из центральной части Европейской России. Вполне вероятно, что существенную роль в его расположении и направленности играет гидрология региона: миграция идет с верховий Волги и Оки вдоль долины Дона в бассейн Северского Донца и дальше на восточное побережье Азовского моря. Долины крупных рек и выступают теми экологическими руслами, которые собирают потоки мигрантов. Причем они важны не только из-за наличия благоприятных мест для отдыха и кормежки, но и являются удобными ландшафтными ориентирами, ведущими кратчайшим путем к Азовскому морю (Грищенко, 1993). Понятно, что такой ход миграции энергетически более выгоден и он будет закрепляться эволюционными механизмами.
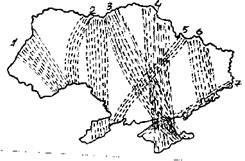
Пролетные пути: Flyways:
1 - Карпатский Carpathian
2 - Бессарабский (Западный) Bessarabian (West)
3 - Днепровский Dnieper
4 - Крымский Crimean
5 - Юго-Западный South-West
6 - Восточный East
7 - Приморский Seaside
Рис. 6. Схема основных пролетных путей осенней миграции птиц на Украине.
Многочисленными исследованиями в последнее время доказано, что сроки и направление миграции птицы определяются особой пространственно-временной генетической программой. Наличие ее уже доказано для 20 видов из 10 семейств (Berthold, 1990a, 1990b). Наблюдаемая в природе картина перелета возникает благодаря взаимодействию условий внешней среды с такой эндогенной программой (подобно этому образуется фенотип на основе генотипа). Вполне возможно, что генетически закреплена информация не только о том, когда и куда, но и где лететь птице, т. е. о расположении пролетных путей.
Итак, резюмируя все вышесказанное, по нашему мнению, упорядоченность структуры миграционного ареала в виде сети пролетных путей различного порядка энергетически выгодна мигрирующим популяциям птиц и развилась в ходе эволюции вслед за возникновением самих миграций.
Борисов А.А. (1975): Климаты СССР в прошлом, настоящем и будущем. Ленинград: ЛГУ. 1-432.
Браунер А.А. (1916): О весеннем пролете аиста. - Орнитол. вестник. 2: 109-119.
Грищенко В.М. (1992): Хiд прильоту шпака в районi Карпат. - Беркут. 1: 78-85.
Грищенко В.Н. (1993): Хорологические особенности осенней миграции птиц на востоке Украины. - Птицы бассейна Северского Донца. Донецк: ДонГУ. 23-25.
Грищенко В.Н. (1994а): Фенологическое картирование в изучении миграций птиц. - Беркут. 3 (1): 30-38.
Грищенко В.Н. (1994б): Фенологiчнi закономiрностi осiнньої мiграцiї птахiв на територiї України. - Дис. ... канд. бiол. наук. Київ. 1-230.
Грищенко В.Н., Серебряков В.В. (1988): Ход весенней миграции чибиса на Украине по данным фенологических наблюдений. - Кулики в СССР: распростр., биология и охрана. М.: Наука. 41-44.
Грищенко В.Н., Серебряков В.В. (1990): Ход весенней миграции черного коршуна на Украине по данным фенологических наблюдений. - Вестн. зоологии. 5: 79-81.
Грищенко В.Н., Серебряков В.В. (1992): Миграции белого аиста на Украине по данным фенологических наблюдений. - Сез. миграции птиц на территории Украины. Киев: Наукова думка. 258-273.
Грищенко В.Н., Серебряков В.В. (1993а): Миграции и зимовки скворца на Украине по данным фенологических наблюдений. - Вестн. зоологии. 3: 59-65.
Грищенко В.Н., Серебряков В.В. (1993б): Миграции серой цапли на Украине по данным фенологических наблюдений. - Бюл. МОИП. Отд. биол. 98 (5): 33-37.
Карри-Линдал К. (1984): Птицы над сушей и морем. Глобальный обзор миграций птиц. - М.: Мысль. 1-204.
Кумари Э.В. (1957): К теории пролетных путей и миграции широким фронтом. - Тр. II Прибалт. орнитол. конфер. М. 4-12.
Лебедева М.И. (1979): Миграции европейского белого аиста по данным кольцевания в СССРю - Миграции птиц Вост. Европы и Сев. Азии. Аистообразные - пластинчатоклювые. М.: Наука. 18-27.
Михеев А.В. (1992): Пролетные пути и широкий фронт пролета птиц. - Успехи соврем. биологии. 112 (2): 298-316.
Полуда А.М. (1983): Некоторые закономерности сезонных миграций птиц в районе Киевского водохранилища. - Вестн. зоологии. 5: 47-53.
Серебряков В.В. (1989): Миграции серого журавля на Украине. - Сообщ. Прибалт. комиссии по изуч. мигр. птиц. Тарту. 21: 122-133.
Bellrose F.C. (1968): Waterfowl migration corridors east of the Rocky Mountains in the United States. - Illinois Natur. Hist. Survey, Urbana, Biol. Notes. 61: 1-24.
Bernis F. (1960): About wintering and migration of the Common Crane in Spain. - Proc. XII Intern. Orn. Congress. Helsinki. 110-117.
Berthold P. (1990a): Spatiotemporal programs and genetics of orientation. - Experientia. 46 (4): 363-371.
Berthold P. (1990b): Genetics of Migration. - In: Gwinner E. (Ed.). Bird Migration. Berlin, Heidelberg: Springer-Verlag. 269-280.
Cameron R.A.D., Cornwallis L., Percival M.J.L., Sinclair A.R.E. (1967): The migration of raptors and storks through the Near East in autumn. - Ibis. 109 (4): 489-501.
Cameron R.A.D., Cornwallis L. (1966): Autumn notes from Azraq, Jordan. - Ibis. 108 (3): 284-287.
Creutz G. (1988): Der Weißstorch. Neue Brehm-Bücherei. 375. Wittenberg Lutherstadt: A. Ziemsen Verlag. 1-236.
Dallinga J.H., Schoenmakers S. (1989): Population changes of the White Stork since the 1850s in relation to food resources. - Weißstorch - White Stork. Proc. 1 Intern. Stork Conserv. Symp. Schriftenreihe des DDA. 10: 231-262.
Geyr von Schweppenburg H. (1931): Zur Terminologie des Vogelzuges. - Proc. VIIth Intern. Orn. Congress. Amsterdam. 333-339.
Heckenroth H. (1968): Beobachtungen über die Rechtwendung des wegziehenden Weißstorchs am Golf von Iskenderun. - Vogelwarte. 24 (3/4): 246-262.
Koch A., Magnus D., Seilkopf H., Baron H., Schüz E. (1966): Der Weißstorch-Zug im Raum Sinai bis Kena in landschaftmorfologischer Sicht. - Vogelwarte. 23 (3): 209-220.
Libbert W. (1936): Der Zug des Kranichs (Grus grus). - J. Ornithol. 84: 297-337.
Lincoln F.C. (1935): The migration of North American birds. - U.S. Dep. of Agriculture Circular. 363: 1-72.
Norrevang A. (1959): Das Zugbild des Kranichs (Grus grus (L.)). - Dansk Ornithol. Forening Tids. 53: 103-109.
Safriel U. (1968): Bird migration at Elat, Israel. - Ibis. 110 (3): 283-320.
Sanden E. (1983): Über den Herbstzug des Kranichs (Grus grus) in Niedersachsen in 1979-1981. - Vogelk. Berichte Niedersachsen. 15 (3): 65-74.
Schulz H. (1988): Weißstorchzug. Ökologie, Gefährdung und Schutz des Weißstorchs in Afrika und Nahost. Königslutter-Lelm. 1-459.
Schüz E. (1968): "Zug-Korridore" von Anatiden in Nordamerika. - Vogelwarte. 25 (1): 108-110.
Schüz E. (1971): Grundriß der Vogelzugskunde. Hamburg und Berlin: P. Parey. 1-390.
Schüz E. (1980): Ungewöhnlicher Zug des Weißstorchs über Zypern. - Vogelwarte. 30 (3): 338-339.
Serebryakov V.V., Grishchenko V.N., Poluda A.M. (1991): The migration of swans, Cygnus spp., in the Ukraine, USSR. - Proc. Third IWRB Intern. Swan Symp., Oxford, 1989. Wildfowl. Supplement No 1: 218-223.