Что такое стволовые клетки
Л.И. Корочкин, доктор медицинских наук, член-корреспондент РАН
Понятие «стволовые клетки» впервые появилось в России еще в начале прошлого века. Тогда великий российский гистолог А.А.Максимов, изучая процесс кроветворения, пришел к выводу об их существовании. Что же это за клетки и каковы их отличительные свойства?
Стволовые клетки могут давать начало любым клеткам организма — и кожным, и нервным, и клеткам крови. Сначала полагали, что во взрослом организме таких клеток нет и существуют они лишь в самом раннем периоде эмбрионального развития. Однако в 70-е годы А.Я.Фриденштейн с соавторами обнаружили стволовые клетки в мезенхиме (строме) «взрослого» костного мозга, в; дальнейшем их стали называть стромальными клетками.
Тогда же появились работы, доказывающие наличие стволовых клеток практически во всех органах взрослых животных и человека. В связи с этим принято разделять стволовые клетки на эмбриональные стволовые клетки (выделяют из эмбрионов на стадии бластоцисты — очень ранней стадии развития, когда еще нет ни тканей, ни закладок органов) и региональные стволовые клетки (выделяют из органов взрослых особей или из органов эмбрионов более поздних стадий), которые сохраняют свойства эмбриональных клеток, о чем свидетельствуют обнаруженные в них эмбриональные белковые маркеры (рис.1).
Стволовые клетки можно выделять и растить в культуре ткани. При этом образуются шарообразные клеточные ассоциаты: скопления эмбриональных клеток называют эмбриоидными телами, а нейральных — нейросферами.
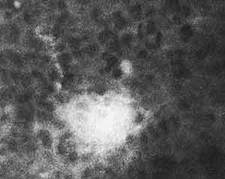
Рис.1. Островки клеток, сохранивших эмбриональные белки (светлые клетки, выявленные с помощью иммунофлюоресцентной техники), в печени взрослой мыши. Препарат получен в 1971 г. в лаборатории Л.И.Корочкина в Новосибирском институте цитологии и генетики СО АН СССР.
Способность давать множество разнообразных клеточных типов (плюрипотентность) делает стволовые клетки важнейшим восстановительным резервом в организме, который используется для замещения дефектов, возникающих в силу тех или иных обстоятельств.
Особое удивление биологов вызвало присутствие стволовых клеток в центральной нервной системе. Как известно, сами нервные клетки утрачивают способность к размножению уже на самой ранней стадии нейральной дифференцировки (стадии нейробласта). А стволовые клетки в ответ на различные поражения нервной ткани начинают делиться с последующей дифференцировкой в нервные и глиальные клетки. Изолированные нейральные стволовые клетки могут превращаться и в другие производные.
Обнаружить стволовые клетки можно с помощью специальных методов. Дело в том, что в «нативных» стволовых клетках и их производных синтезируются специфические белки, которые выявляются с помощью иммуногистохимической техники. На каждый белок получают антитела, которые метят флюоресцирующим красителем. Такой реагент выявляет белки, присутствующие в стволовых клетках на разных стадиях развития. Так, нейральные стволовые клетки содержат белок нестин (рис.2). Когда они вступают на путь специализации, в них появляется новый белок — виментин. Если клетки развиваются в нейральном направлении, то синтезируются соответствующие маркирующие белки — нейрофиламентные, b3-тубулин, энолаза и др. Когда клетки специализируются как вспомогательные, глиальные, появляются другие маркеры, например глиальный фибриллярный кислый белок, белок S-100 и др.
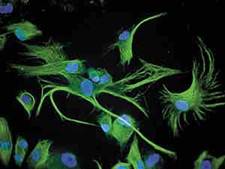
Рис.2. Культура стромальных стволовых клеток человека.
Зеленым флюоресцирует цитоплазма, содержащая нестин, синим — ядерный материал.
Модель для анализа роли генов в дифференцировке
Способность любых стволовых клеток давать разные клеточные типы делает их весьма удобной системой для изучения молекулярно-генетических событий, обусловливающих специфическую дифференцировку клеток. Действительно, изолировав стволовые клетки в чистом виде, можно затем анализировать функции генов, ответственных за последовательные этапы дифференцировки.
Оказалось, в частности, что время последовательного включения генов, контролирующих развитие, совпадает и в постимплантационных зародышах, и в культуре эмбриоидных тел. Значит, стволовые клетки — действительно хорошая экспериментальная модель для изучения молекулярных механизмов клеточной специализации.
Анализ культур стволовых клеток с помощью молекулярно-генетического микроэррэй-метода (microarray), оценивающего количество функционально активных генов, показал, что в одном клоне мезенхимных стволовых клеток синтезируется по крайней мере 1200 матричных РНК (мРНК). В разных стволовых клетках присутствует сходный набор заранее синтезированных мРНК (копий многих генов), но есть и специфические РНК. При этом удалось выяснить, что в стромальных стволовых клетках взрослой гематогенной (кровеобразующей) ткани содержится практически весь набор мРНК, которые функционируют в зародышевых листках и на стадии органогенеза. Идентифицированы также мРНК ключевых генов, регулирующих созревание клеток всех зародышевых листков: мезенхимального и мезодермального происхождения, а также энто- и эктодермы. Большинство мРНК регуляторных генов присутствует уже в яйцеклетке и зародышевых клетках.
Следовательно, в стволовых клетках проявляется общий принцип онтогенеза — работа генов с «опережением», т.е. синтез тех мРНК, которые понадобятся на значительно более поздних стадиях развития.
Гены-господа, проблема дифференцировки и стволовые клетки
Многочисленные данные, полученные в ходе изучения стволовых клеток, позволили уточнить организацию соответствующих генных сетей (в условиях целостного организма это иногда не так-то просто). В частности, можно выявить пути взаимодействия так называемых генов-господ и генов-рабов. Господами называют ключевые гены, от которых зависит специфика развития данной ткани или органа, рабами — каскады структурных генов (запускаемые генами-господами), обеспечивающих синтез тканеспецифических белков и соответственно формирование того или иного органа или ткани.
Использование стволовых клеток в биологии развития позволило подтвердить существование генов-господ, запускающих каскады генов, от которых зависит специализация целых органов, зародышевых листков и отдельных типов клеток. Эта универсальная закономерность присуща всем животным. Так, у дрозофилы есть ген eyeless (безглазости), который обусловливает развитие глаза. Если его заставить работать в необычном месте, то глаза могут появиться на брюхе, на лапках, на крыле и в любом другом месте (рис.3). Сходный ген Pax6 есть и у млекопитающих. Введенный в геном дрозофилы, он дает тот же эффект, что и собственный ген хозяина. Все это свидетельствует об универсальности эффекта генов-господ.
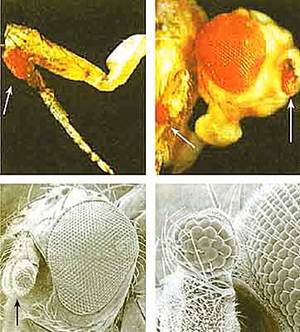
Рис.3. Развитие глаза в необычных местах на теле дрозофилы: а, б — на лапке, в, г — на антенне.
Ген pdf-1 выполняет роль триггера, запускающего развитие поджелудочной железы; ген НОХ-11 отвечает за развитие селезенки, ген Crypto — за развитие сердца, мутации гена НОХD13 приводят к полидактилии верхних и нижних конечностей у человека. Известны гены-господа и для отдельных зародышевых листков. Так, мутация гена casanova блокирует развитие всей энтодермы, а генов Brachiury и zeta-globin — мезодермы.
Наконец, по сигналу соответствующих генов-господ формируются специализированные ткани и типы клеток. Например, ген Wn17 инициирует созревание альвеолярного эпителия. В нашей лаборатории совместно с лабораторией В.Тарабыкина (университет Геттингена) открыта новая группа нейрогенов, необходимых для образования нейронов пятого-шестого слоев коры головного мозга.
Возможно, определенную регулирующую роль в дифференцировке стволовых клеток играют короткие повторяющиеся последовательности, микро- или минисателлитные. Так, О.В.Подгорная (Санкт-Петербург) обнаружила наличие белков, специфическое связывание которых с тандемными повторами определяет особенности трехмерной организации хроматина. Как известно, от этой организации зависит специфика работы генов. Значит, состояние системы повторяющихся последовательностей (их недорепликация, диминуция или гиперрепликация) может играть важную роль в дифференцировке стволовых клеток.
Сегодня очевидно, что индивидуальное развитие регулируется иерархически организованной системой генных ансамблей (сетей). Понять особенности такой регуляции помогают стволовые клетки. В связи с этим большой интерес представляет реконструкция органных структур in vitro на основе стволовых клеток. Так, М.Томоока с соавторами получили из стволовых нервных клеток структуры, подобные нервной трубке; сходные опыты с диссоциированными клетками гиппокампа поставил в Институте мозга РАМН И.В.Викторов. Предпринимаются также попытки выращивать клетки в специальных колонках для получения органоподобных структур и использования их в клинике. Такие исследования весьма перспективны и для решения фундаментальных задач, и для практического использования в генной и клеточной терапии.
Стволовые и камбиальные клетки
Давно известно, что почти каждая ткань в организме имеет запас так называемых камбиальных клеток, которые пополняют ее клеточный состав, постоянно тающий от функциональных перегрузок или болезней. При столь пристальном внимании к стволовым клеткам немудрено, что камбиальные клетки преданы забвению. О них вообще как-то стали забывать. А между тем камбиальные клетки — непосредственный участник восстановительных процессов в тканях. Наглядный тому пример — клетки росткового слоя кожи, пополняющие постоянно расходуемый запас зрелых, уже не делящихся клеток кожного покрова. Более того, до открытия стволовых клеток речь шла только о таком способе репарации. В нервной ткани камбиальных клеток, способных размножаться, нет (о глии речь не идет). Но там сохраняется резерв молодых клеток — нейробластов, которые благодаря своей дифференцировке восполняют различные дефекты, сохраняя тем самым функциональную дееспособность соответствующего отдела мозга или периферической нервной системы.
Каковы же взаимоотношения стволовых и камбиальных клеток? Возможны ли их взаимопревращения, может ли региональная стволовая или прогениторная клетка (клетка-предшественник) дать начало камбиальной и наоборот, происходит ли этот процесс в организме, каково его значение для нормального течения восстановительных процессов и каков (если он существует) его молекулярно-генетический механизм? Решение этих вопросов имеет важное не только фундаментальное, но и практическое значение. Изучение стволовых клеток в разных экспериментальных условиях, бесспорно, поможет найти ответы и позволит представить в новом свете тонкие механизмы восстановительных процессов, протекающих в организме. Такие работы уже начаты, в частности на стволовых клетках эпителиального покрова кожи. Результаты противоречивы и дают повод для дискуссий.
При этом следует учитывать, что в самую начальную фазу дифференцировки с разной степенью эффективности включается несколько программ, и судьба клеток еще однозначно не решена. Например, в развивающемся нейробласте, дифференцирующемся в катехоламинэргическом направлении, синтезируются не только мРНК для компонентов катехоламинэргической системы, но и мРНК для компонентов холинэргической системы. Если в определенный момент развития сменить катехоламинэргическую мишень, иннервируемую данной клеткой, на холинэргическую, то ранее более интенсивный синтез «катехоламинэргических» РНК начнет тормозиться и возобладает синтез «холинэргических» РНК. В результате произойдет как бы перепрограммирование клетки на новый путь развития.
Стволовые клетки, встречающиеся в шиповатом слое эпидермиса кожи, чьи клетки уже не делятся и активно специализируются, как раз и могут быть «мигрантами» из очага стволовых клеток. Мне доводилось встречать такие клетки в дифференцирующейся автономной нервной системе эмбрионов человека. Иными словами, ситуация с «превращениями» стволовых клеток и их взаимоотношениями с камбиальными клетками далеко не так проста, как это может показаться на первый взгляд.
Изменились ли представления о клеточной дифференцировке с открытием стволовых клеток?
Вопреки утверждению некоторых авторов — пока нет. Во-первых, дифференцировка любых стволовых клеток происходит по законам, сформулированным для клеточной дифференцировки вообще. В этом и заключается ценность стволовых клеток как модельной системы. Во-вторых, клетки, в том числе и стволовые, начав дифференцировку, утрачивают способность к делению, по крайней мере на конечных стадиях. И, наконец, изучение поведения стволовых клеток не поколебало представлений о стабильности и необратимости клеточной дифференцировки: из фиброцита, плазматической или из париетальной клетки желудка никогда не получится нейрон, а из нейрона не возникнет кожная клетка. Тезис, что стволовая клетка способна к разного рода трансформациям, никак не нарушает это правило, а лишь демонстрирует мультипотентность, свойственную ранним эмбриональным клеткам. На стадии терминальной дифференцировки клетка обретает стабильное состояние и теряет способность к делению и разного рода превращениям.
Что же нового дало открытие стволовых клеток во взрослом организме? Оно изменило наши представления об организации тканей и о механизмах протекающих в них восстановительных процессов. Был сделан новый и очень важный вывод: эмбриональные клетки с высоким потенциалом к развитию сохраняются и во взрослом организме. Более того, они составляют важнейшее звено в цепи репаративных процессов, о чем ранее не подозревали. Так, описывая в 70-е годы эмбриональные клетки в печени взрослой мыши (в книге «Взаимодействие генов в развитии». М., 1977), я не предполагал, что они обладают столь высоким потенциалом к развитию и принимают активное участие в репарации.
Открытие стволовых клеток повлекло необходимость замены существовавшей до сих пор схемы репаративных процессов в тканях:
на новую схему, отражающую существование стволовых клеток во взрослом организме:
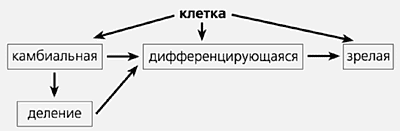
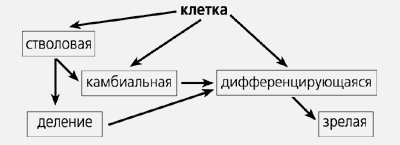
В ходе клеточного деления из стволовых клеток возникают материнская и дочерняя клетки. Материнские используются для самоподдержания популяции, а дочерние либо «выходят» в камбиальную клетку, либо непосредственно в дифференцировку. Стволовая клетка сохраняет свойства ранних эмбриональных клеток — плюрипотентность, а камбиальная эту способность утрачивает и производит лишь региональные структуры.
Таким образом, в изучении восстановительных процессов сделан большой шаг вперед. Но предстоит еще очень много сделать, чтобы познать тонкие механизмы поведения стволовых клеток и найти возможность использовать эти знания в клинической практике.
Ситуация в данном случае непростая. Она осложняется еще и присутствием маркеров, которые специфически «метят» стволовые клетки и их производные. В частности, в базальном, ростовом слое эпителия кожи имеется стволовых клеток 10%, а белок b1-интегрин, специфический маркер этих клеток, содержится у 40% клеток этого слоя. В связи с этим сообщения о взаимопревращениях и трансформациях кожных (да и других тоже!) стволовых клеток требуют серьезной экспериментальной проверки.
Трансдетерминация и трансдифференцировка
В связи с необычайно широким потенциалом стволовых клеток возникает путаница с понятиями трансдетерминации и трансдифференцировки. В результате принятые в гистологии и эмбриологии терминологические правила размываются и возникает почва для бесплодных дискуссий и спекуляций.
Действительно, если трансформацию стволовых клеток в разных направлениях обозначить как трансдифференцировку (а некоторые авторы позволяют себе такую вольность), будут необоснованно разрушены представления о стабильности и необратимости дифференцировки, что ведет к невообразимой путанице. На самом деле нет никаких оснований ниспровергать существующие взгляды. Совершенно очевидно, что клетка, потерявшая способность к делению и вступившая на определенный путь развития (например, нейробласта), не может дать начало другим производным. Добиться репрограммирования ядра не так-то просто. Даже его пересадка в другую цитоплазму (в частности, при получении гетерокарионов или в опытах с пересадкой ядер), и то не всегда успешна.
Зарегистрированные случаи трансформации стволовых клеток относятся к другому событию — трансдетерминации. Процесс этот давно известен в экспериментальной эмбриологии благодаря работам выдающегося швейцарского эмбриолога и генетика Эрнста Хадорна. Описанное в ряде работ «превращение» глиальной клетки в нейрон объясняется, видимо, гетерогенностью популяции глиоцитов, т.е. некоторые из них могут сохранять свойства камбиальности, а порою и «стволовости». В таком случае обнаруженный феномен удивления не вызывает. Например, показано, что клетки так называемой радиальной глии, которая на ранних этапах онтогенеза служит субстратом для миграции дифференцирующихся нервных клеток, становятся нейронами. Однако потом выяснилось, что на самом деле популяция клеток радиальной глии гетерогенна: часть клеток содержит нейральные маркеры (они впоследствии становятся нервными), а часть — глиальные (такие и становятся глиальными). Иными словами, несмотря на то, что все клетки радиальной глии вначале выполняют одну и ту же временную функцию, они уже детерминированы к развитию в разных направлениях. Значит, обнаруженный феномен их трансформации — не трансдифференцировка, а трансдетерминация.
Как стволовые клетки поддерживают свою «молодость»
Одна из важнейших общебиологических проблем, решить которую помогут стволовые клетки, — генетический механизм поддержания детерминированного состояния в ходе деления клеток и выхода их в дифференцировку. Всерьез ее поставил еще Э.Хадорн в 50-е годы прошлого века, но до сих пор она не решена. Недавно удалось пролить некоторый свет на молекулярно-генетические события при переходе клетки из детерминированного состояния в дифференцировку. Наша соотечественница Наталья Тулина, работающая в США, заметила, что для такого перехода очень важно взаимоотношение стволовых клеток с клетками-«нишами», к которым они «прилежат». Так, в семенниках дрозофилы соматические клетки «хаба», формирующие нишу стволовых клеток, содержат белок UPD, который, в свою очередь, активирует так называемый сигнальный каскад Jak-STAT. Усиленный синтез UPD в клетках апикального района семенников приводит к росту и репродуктивных, и стволовых клеток семенника. Для поддержания обоих типов клеток необходимо участие компонентов Jak-STAT сигнального каскада, киназы НОР и транскрипционного активатора STAT92E. Активацию всего комплекса белков запускает UPD, который клетки-«ниши» передают стволовым клеткам. Разрыв связи между ними обусловливает начало дифференцировки стволовых клеток (рис.4). Насколько универсален этот механизм, предстоит еще выяснить.
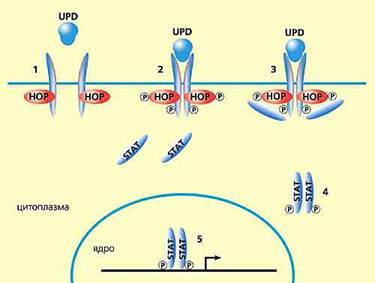
Рис.4. Схема поддержания «стволовости» клеток в семенниках дрозофилы по данным Натальи Тулиной.
1, 2 и 3 — стадии взаимодействия с комплексом мембранных белков,
4 — активация сигнального белка STAT,
5 — активация генов стволовой клетки.
Проблемы генной и клеточной терапии
Плюри- и мультипотентность стволовых клеток делает их идеальным материалом для трансплантационных методов клеточной и генной терапии. Наряду с региональными стволовыми клетками, которые при повреждении тканей соответствующего органа мигрируют к зоне повреждения, делятся и дифференцируются, образуя в этом месте новую ткань, существует и «центральный склад запчастей» — стромальные клетки костного мозга. Эти клетки универсальны. Они, видимо, поступают с кровотоком в поврежденный орган или ткань и там под влиянием различных сигнальных веществ продуцируют взамен погибших нужные клетки (хотя полученные многочисленные данные такого рода нередко критикуются и требуют дополнительной проверки).
В частности, установлено, что инъекция экспериментальным животным стромальных клеток костного мозга в зону повреждения сердечной мышцы устраняет явления постинфарктной сердечной недостаточности. А стромальные клетки, введенные свиньям с экспериментальным инфарктом, уже через восемь недель полностью перерождаются в клетки сердечной мышцы, восстанавливая ее функцию. Результаты такого лечения инфаркта впечатляющи. По данным Американского кардиологического общества, за 2000 г. у крыс с искусственно вызванным инфарктом 90% стромальных клеток костного мозга, введенных в область сердца, трансформировались в клетки сердечной мышцы.
Японские биологи в лабораторных условиях получили клетки сердечной мышцы из стромальных клеток костного мозга мышей. В культуру стромальных клеток добавляли 5-азацитидин, и они начинали превращаться в клетки сердечной мышцы. Такая клеточная терапия весьма перспективна для восстановления сердечной мышцы после инфаркта, поскольку для нее используются собственные стромальные клетки. Они не отторгаются, и, кроме того, при введении взрослых стволовых клеток исключена вероятность их злокачественного перерождения.
Широко применяется терапия стромальными клетками в ортопедии. Это связано с существованием особых белков, так называемых ВМР (костные морфогенетические белки), которые индуцируют дифференцировку стромальных клеток в остеобласты (клетки костной ткани). Клинические испытания в этом направлении дали многообещающие результаты. Например, в США 91-летней пациентке с незаживающим в течение 13 лет переломом вживили специальную коллагеновую пластинку с нанесенными на нее ВМР. Поступающие в зону перелома стромальные клетки «притягивались» к пластинке и под влиянием ВМР превращались в остеобласты. Через восемь месяцев после установки такой пластинки сломанная кость у больной восстановилась. Сейчас в США проходят испытания и скоро начнут применяться в клинике специальные пористые губки, наполненные одновременно и стромальными клетками и нужными индукторами, направляющими развитие клеток по требуемому пути.
Большое значение придают стволовым клеткам (в частности, стромальным) при лечении различных нейродегенеративных и неврологических заболеваний — паркинсонизма, болезни Альцгеймера, хореи Гентингтона, мозжечковых атаксий, рассеянного склероза и др. Группа неврологов из Американского национального института неврологических заболеваний и Стэнфордского университета обнаружила, что стромальные стволовые клетки костного мозга могут дифференцироваться в нейральном направлении. Значит, костный мозг человека можно использовать как источник стволовых клеток для восстановления поврежденных тканей в головном мозгу. При этом, видимо, возможен не только заместительный, но и трофический эффект трансплантата (это предположение основано на том, что положительное действие трансплантата проявляется через две недели, а эффект замещения возможен лишь спустя три месяца). Следовательно, пациент может стать собственным донором, что предотвратит реакцию иммунологической несовместимости тканей.
Группа американских ученых под руководством Е.Мизей показала, что стволовые клетки, куда бы их ни имплантировали, способны достигать поврежденного места, в частности мозга, и обеспечивать там восстановительные процессы. Так, после внутривенного введения взрослым мышам стромальных стволовых клеток во многих областях мозга (включая неокортекс, гиппокамп, таламус, ствол мозга и мозжечок) были обнаружены различные нейральные производные. Впрочем, литературные данные по этой проблеме весьма противоречивы. Однако если к культуре стромальных стволовых клеток добавить ретиноевую кислоту, в них обнаруживаются нейральные маркеры (рис.5). Такие клеточные культуры харьковские хирурги небезуспешно применяли для лечения болезни Паркинсона, вводя их в область полосатого тела (рис.6).
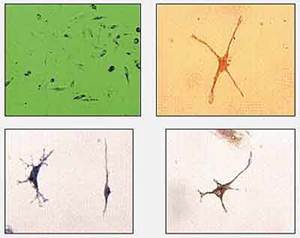
Рис.5. Дифференцировка нервноклеточных элементов в культуре стромальных стволовых клеток человека под влиянием ретиноевой кислоты: а — исходная культура, б — окраска на нейрофиламенты, в, г — окраска на b3-тубулин.

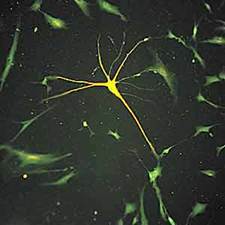
Рис.6. Иммунофлюоресцентная окраска на нейроспецифические маркеры клеток, дифференцирующихся в нейральном направлении, в культуре стромальных стволовых клеток костного мозга человека. Такие клетки использовали харьковские хирурги для лечения болезни Паркинсона.
Весьма перспективны также попытки использовать стволовые клетки пуповины и плаценты в клинике. В целом для успешной пересадки стволовых клеток, независимо от области применения, очень важно научиться сохранять их жизнеспособность. Ее можно повысить, если в геном пересаживаемых нейронов вводить гены ростовых нейротрофических факторов, которые служат защитой от апоптоза. Такие попытки ведутся в различных лабораториях США и Европы.
Больших успехов в изучении и практическом использовании стволовых клеток добились и отечественные исследователи. Специалисты из Института акушерства, гинекологии и перинатологии РАМН выделили региональные нейральные стволовые клетки и впервые получили их подробную иммуногистохимическую характеристику, в том числе на проточном флюориметре. В опытах с пересадкой стволовых нейральных клеток человека в мозг крыс показана их приживляемость, миграция на достаточно большие расстояния (несколько миллиметров) и способность к дифференцировке, которая в значительной степени определялась микроокружением трансплантата. Например, при пересадке нейральных клеток человека в область мозжечка крысы, где расположены клетки Пуркинье, они развиваются в направлении именно этого типа клеток. Об этом свидетельствует синтез в них белка калбиндина, специфического продукта клеток Пуркинье.
Отечественные биологи (Институт биологии гена РАН, Харьковский институт криобиологии и фирма «Виола») впервые разработали оригинальную методику индукции в культуре: стромальные стволовые клетки дифференцировались в направлении клеток, похожих на клетки островков Лангерганса, содержащих инсулин. Наличие этого белка в них определяли с помощью современных методов молекулярной биологии и цитологии. Самое интересное, что в культуре эти клетки формируют структуры, напоминающие островки Лангерганса. Не исключено, что в далекой перспективе они станут незаменимы для лечения диабета.
Интересную совместную работу провели сотрудники трех академических научных учреждений — Института биологии гена, Института биологии развития и Института молекулярной биологии. При пересадке кусочков эмбриональной нервной ткани дрозофилы в мозг крысы заметили, что вокруг трансплантата не формируется рубцовая ткань. Оставалось выяснить, за счет чего это происходит. С помощью достаточно тонких экспериментов удалось установить, что образованию рубца препятствуют белки теплового шока, которые синтезируются в клетках дрозофилы при температуре тела млекопитающих. Значит, добавление ксенотрансплантата (ткани дрозофилы) к эмбриональной нервной ткани крысы спасает аллотрансплантат от нашествия рубцовой ткани (рис.7). Так появилась возможность использовать белки теплового шока в клеточной и генной терапии различных заболеваний.
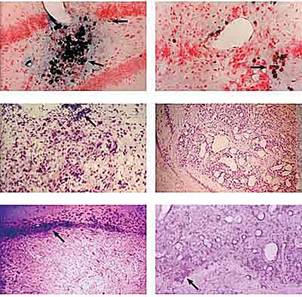
Рис.7. Трансплантация клеток дрозофилы в мозг крысы (ксенотрансплантация). а, б — меченые клетки дрозофилы (стрелки) в мозге крысы; в — аллотрансплантат эмбриональной нервной ткани крысы в сочетании с ксенотрансплантатом (стрелка); г — стимуляция роста кровеносных сосудов ксенотрансплантатом; д — рубцовая ткань (стрелка) на границе аллотрансплантат/ткань хозяина; е — добавление ксенотрансплантата к аллотрансплантату блокирует образование рубца.
Подобные исследования позволят создавать генно-инженерные конструкции для трансформации стволовых клеток, предназначенных для пересадки. Эти структуры помогут лучшему приживлению трансплантата, повысят его жизнеспособность и специализацию составляющих его клеток.
Необходимо сравнить и внимательно проанализировать результаты трансплантации стволовых клеток в виде цельных или диссоциированных на клетки нейросфер и разработать соответствующий протокол для клинического использования.
Впрочем, нельзя не сказать о том, что из достаточно авторитетных лабораторий раздаются весьма скептические отклики на такие работы и предупреждения о необходимости осторожно интерпретировать полученные данные. Приводятся факты, свидетельствующие, что стволовые клетки не дифференцируются после их трансплантации, а сливаются со специализированными клетками хозяина, создавая видимость собственной дифференцировки. Некоторые авторы считают, что стромальные клетки костного мозга способны превращаться только в клетки хряща и кости, а при их инъекции реципиенту оседать там, откуда «пришли», т.е. в костном мозгу, в связи с чем перспективы их использования в клеточной терапии ставятся под сомнение. Очевидно, необходимы дополнительные серьезные исследования для ответа на поставленные вопросы и выдвинутые возражения.
1. Корочкин Л.И. Биология индивидуального развития. М., 2002.
2. Репин В.С., Ржанинова А.А., Шаменков Д.А. Эмбриональные стволовые клетки: фундаментальная биология и медицина. М., 2002.
3. Репин В.С., Сухих Г.Т. Медицинская клеточная биология. М., 1998.
4. Онтогенез. 2003. Т.34. №3. Выпуск целиком посвящен проблеме стволовых клеток.