Поведение чайки
Поведение чайки
Полевое и лабораторное изучение
реферат

Бабицкий Мирослав, II курс, 6 группа, биологический факультет БГУ.
Обыкновенная, или озерная, чайка (L. ridibundus) — обычная и почти всюду наиболее многочисленная из наших чаек. По размерам она несколько мельче сизой чайки, весит 250—400 г. Телосложение ее стройное, полет легкий, маневренный, обычно довольно размеренный. Окрашена она в беловатые тона снизу, серовато-дымчатые сверху, с темно-коричневой головой и черными концами крыльев.
Озерная чайка широко распространена в средней части Евразийского материка. Обитает по разнообразным внутренним водоемам, на пролете и зимовках держится также и по морским побережьям.
Весной озерные чайки прилетают рано, когда начинают вскрываться водоемы и еще не полностью сойдет снег. В зависимости от географического положения местности это наблюдается от конца февраля до конца апреля. Первое время после прилета птицы кочуют поблизости от гнездовых мест и к устройству гнезд приступают сравнительно поздно, после спада талых вод.
В размножении участвуют птицы в возрасте около двух лет и реже в годовалом возрасте. Гнездятся колониями, размер которых весьма различен, от немногих до нескольких тысяч пар. Иногда колонии смешаны с другими видами чаек и крачек. Гнездовыми местами служат преимущественно стоячие и медленно текущие водоемы — озера, болота, речные заводи и протоки, окруженные растительностью или имеющие плавни. Колонии обычно располагаются на одних и тех же труднодоступных местах — на плавнях, топких берегах и т, д.
В апреле - мае, а на севере даже в начале июня самка откладывает в гнездо 3, реже 2 или 4 яйца грязно-зеленоватого цвета с серыми и бурыми пятнами. Размеры яиц: 51—67 X 36—41 мм. Насиживают обе птицы на протяжении 22— 24 дней.
Вылупившиеся птенцы через 12 —16 часов уже могут стоять. С этого же времени, а иногда и через сутки родители начинают их кормить, а до этого они существуют за счет остатков желточного мешка. Кормят взрослые птенцов 4—5 раз в день отрыжкой, выкидывая ее им в рот. Уже в двухдневном возрасте птенцы начинают делать попытки склевывать отрыгнутый перед ними корм.
Примерно в возрасте 10 дней молодые чайки перебираются от гнезда в заросли густой растительности, держась при этом выводком. «Чужого» птенца, пробегающего мимо гнезда, а тем более пытающегося в него залезть или присоединиться к выводку, старая чайка обычно убивает ударом клюва в голову. При этом страдают как мелкие пуховички, так и крупные, почти оперившиеся птенцы. На территории гнездовой колонии чаек всегда можно встретить трупы птенцов. В возрасте 18—20 дней птенцы начинают бродить самостоятельно, и взрослые перестают относиться враждебно к «посторонним» молодым чайкам.
В пятинедельном возрасте молодые оперяются и начинают летать, но совсем летными становятся в шестинедельном возрасте. В средней полосе в середине июля начинают отлетать из гнездовой колонии самцы, дней через десять за ними следуют самки, а в начале августа и молодые. Период размножения кончается, и начинаются послегнездовые кочевки, постепенно переходящие в осенний перелет. Последний протекает в сентябре, затягиваясь на юге до зимы.
Питаются озерные чайки в основном животными кормами: водными и наземными насекомыми, мышевидными грызунами, рыбами, лягушками, дождевыми червями. Рыб чайки ловят только с поверхности воды, и главным образом больных особей. Птенцов выкармливают в основном насекомыми и дождевыми червями. Корм собирают в ближайших окрестностях гнездовой колонии и, как исключение, летают на богатые кормовые угодья на расстояние до 20 км от гнезд.
Вылавливая вредных грызунов и насекомых, чайки приносят большую пользу сельскому хозяйству.
Тихо сидя в укрытии поблизости от гнезда, можно наблюдать за кормлением птенцов чаек. Взрослая птица опускает голову с клювом, направленным вниз, перед семидневным птенцом. Если после предыдущего кормления прошло достаточно времени, птенец совершает сложное координированное движение - клюет клюв взрослой птицы и тянет его вниз. После нескольких таких клевков взрослая птица отрыгивает частично переваренную пищу. Таким образом, клевки птенца, по-видимому, являются формой выпрашивания корма. Потом птенец клюет рыбу, отрывая и заглатывая куски. Следовательно, клевки - это также и способ поглощения пищи. Когда птенцы в гнезде уже наелись досыта, птица съедает остатки корма.
Дальнейшие наблюдения позволили выявить ряд более сложных моментов во взаимодействии птенцов и взрослой птицы. Если обычное опускание клюва перед глазами птенцов не вызывает у них пищевой реакции, птица начинает слегка покачивать клювом из стороны в сторону. Такое движение обычно стимулирует клевки. Отрыгнув пищу в гнездо, птица дожидается, пока птенцы начнут кормиться. Если этого не происходит, взрослая птица опять опускает клюв, как бы указывая на пищу. После этого птенцы обычно начинают клевать. Если же птенцы и теперь не реагируют на пищу, птица берет ее в клюв и держит перед ними. Как только птенцы начинают клевать корм, птица сразу отпускает его.
В связи с этими внешне простыми реакциями клевания возникает ряд вопросов относительно возможной роли индивидуального опыта в развитии пищевого поведения. Как может птенец касаться своим клювом клюва взрослой птицы при выпрашивании корма и отрывать куски от принесенной пищи, если обе формы клевка в своей основе столь сходны? Почему птенец поворачивает голову в сторону при клевке во время выпрашивания, но не делает этого, когда клюет пищу? Нужна ли птенцу практика, чтобы добиться точности и координированности клевка? Почему голодный птенец клюет, а сытый - нет? Почему клевки птенца не бывают нацелены на красные лапы взрослой птицы или на другие окружающие предметы? Как птенец распознаёт пищу?

В поисках ответов на эти и многие другие вопросы группа Хейлмена занималась экспериментальным изучением поведения птенцов чайки с момента их вылупления и до достижения недельного возраста. К седьмому дню жизни формирование пищевых реакций у птенцов практически завершается. Кроме того, ограничивая исследование таким коротким периодом, можно более четко контролировать отдельные элементы онтогенеза и оценивать их вклад в поведение. Как это часто бывает, исследование породило больше вопросов, чем разрешило, однако было получено много новых сведений. Рассмотрим сначала развитие точности клевков. Для изучения этой проблемы была схематически изображена голова взрослой птицы на маленьких карточках.
Карточка, прикрепленная к подвижному стержню, могла перемещаться в горизонтальном направлении перед глазами птенца. Собранные в гнездах яйца инкубировались в темноте, так что птенцы до начала эксперимента не получали никаких зрительных стимулов. После вылупления в первый же день каждому птенцу предъявляли подвижную двумерную модель головы взрослой птицы и позволяли ему клюнуть ее 10-15 раз. Место каждого клевка отмечалось на модели точкой.
Слева на рисунке - результат эксперимента, проведенного с только что вылупившимся птенцом, справа-с тем же птенцом через два дня.
После эксперимента, предварительно пометив птенца, возвращали его в гнездо и брали взамен яйцо, из которого были слышны писки, т.е. яйцо с эмбрионом, близким к вылуплению. Меченый птенец, таким образом, воспитывался в естественных условиях приемными родителями. На 1-й, 3-й и 5-й дни после вылупления половину меченых птенцов тестировали вновь. Поведение остальных птенцов исследовали на 2-й, 4-й и 6-й дни. После каждого эксперимента птенцов опять возвращали в гнездо.
Опыты показали, что сразу после вылупления птенцов в среднем только треть их клевков достигает цели. На следующий день точность уже превышает 50%, а через два дня после вылупления достигает величины более 75% и в дальнейшем не меняется. Анализ данных для типичного птенца показывает, что разброс клевков, особенно горизонтальный, значительно уменьшается с возрастом. Так же происходит столь быстрое повышение точности клевков? Для выяснения этого вопроса был разработан более сложный эксперимент с использованием двух контрольных групп птенцов, выращенных в естественных условиях. Чтобы птенцы экспериментальных групп не могли приобрести опыта зрительной координации клевания, их содержали в темных брудерах. Птенцов одной из этих групп кормили искусственно. Птенцы второй группы не получали пищи в течение двух дней: они существовали за счет обильных жировых запасов. Для того чтобы выяснить, влияет ли на точность клевков нормальная активность, связанная с вылуплением, птенцов третьей группы извлекали из яиц сразу после появления эмбриональных писков и помещали в инкубатор.
В различные сроки после вылупления птенцам предъявляли модель головы взрослой птицы. Поведение птенцов регистрировалось на кинопленке, анализ которой позволял определять процент точных клевков. Во всех пяти группах точность с возрастом увеличивалась, но только в двух контрольных группах она достигала нормального уровня (свыше 75%). В отличие от зрительной депривации, оказывавшей заметное влияние на точность клевания, исключение нормальной активности, связанной с вылуплением, практически не влияло на результаты тестирования.
Наиболее осторожной интерпретацией этих результатов мог бы быть следующий вывод: зрительный опыт необходим для достижения максимальной точности клевания, но некоторое повышение точности происходит и в условиях зрительной депривации. В последнем случае улучшение может быть результатом совершенствования позных реакций. Здесь тоже возможно влияние индивидуального опыта, так как весьма вероятно, что позные реакции эффективно отрабатываются у птенцов в условиях темного инкубатора.
Как научается птенец выбирать правильное расстояние для точного клевка? Наблюдения над только что вылупившимися птенцами позволяют предположить, что здесь действует механизм саморегуляции, основанный на восприятии расстояния. Если неопытный птенец встанет слишком близко к модели, то удар оказывается настолько сильным, что он может отбросить птенца назад на 2-3 см. И наоборот, когда птенец находится слишком далеко от цели, он не достает до нее при клевке и падает вперед на 4-5 см. Старшие птенцы редко допускают такие серьезные ошибки; можно думать, что с возрастом они методом проб и ошибок научаются выбирать наилучшее расстояние.
Исследователи нередко рассматривали пищевую мотивацию (голод) как результат научения. Эксперименты позволяют предполагать, что по крайней мере в основе голода лежит нечто врожденное. Ряд опытов показал, что у накормленных досыта птенцов, как и следовало ожидать, частота клевков возрастает с увеличением интервала времени после кормления. Однако такая же картина наблюдалась и у птенцов, которым мы ни разу не доказали случая испытать чувство голода. Птенцов, вылупившихся в темном инкубаторе, искусственно кормили до насыщения на вторые сутки жизни, а затем тестировали на свету с моделями головы. Через час после кормления средняя частота клевков составляла у них 3,1 в минуту, а через два часа-5,1 (разница статистически достоверна).
Детали движений при клевке во время пищевой реакции анализировались с помощью высокоскоростной киносъемки. У птенца в возрасте нескольких дней пищевая реакция, к этому времени уже вполне сформированная, включает четыре основных компонента: 1) открывание и последующее закрывание клюва; 2) движение головы вверх и вперед по направлению к голове взрослой птицы, а затем вниз и назад; 3) поворот головы в сторону в расчете на то, чтобы захватить клювом клюв взрослой птицы, и обратный поворот головы; 4) легкий толчок ногами вверх и вперед (см. рис. 114). Покадровый анализ фильма позволил выявить значительную изменчивость временных отношений между этими компонентами, как у каждого отдельного птенца, так и у различных птенцов. С возрастом эта изменчивость несколько уменьшается, что, вероятно, связано с улучшением координации движений. Среди интересных деталей, выявленных при анализе кинолент, можно отметить тот факт, что у птенцов, выросших в естественных условиях, поворот головы в сторону наблюдался с возрастом все чаще. Чтобы выяснить, насколько это зависит от зрительного опыта, мы проанализировали фильмы, заснятые во время тестов на точность клевания во всех пяти группах птенцов. Оказалось, что у птенцов, не имевших опыта клевания, сколько-нибудь заметный поворот головы наблюдался редко. У птенцов, выросших в гнезде, в первые сутки жизни поворотов головы не бывает, но затем эта реакция появляется и быстро совершенствуется. Не известно, какова конкретная роль индивидуального опыта в таком развитии поведения, но снятые фильмы дают основание для некоторых предположений. Иногда, когда птенец клюет с раскрытым клювом, клюв взрослой птицы, расположенный не строго вертикально, может оказаться между челюстями птенца. Тогда движение головы птенца по инерции вперед поневоле приводит к ее повороту в сторону. Возможно, именно так птенец научается поворачивать голову, чтобы схватить клюв взрослой птицы.
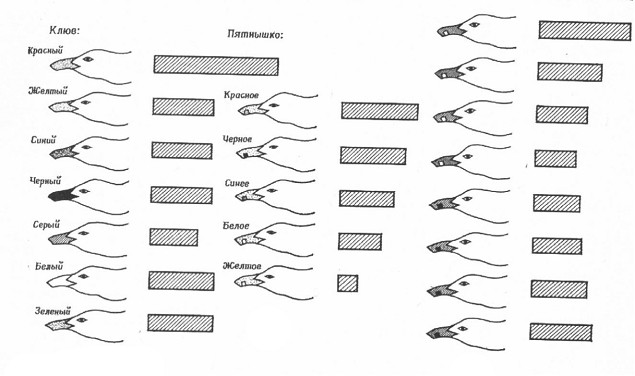
Один из самых интересных вопросов, связанных с пищевым поведением птенцов - это вопрос о том, как птенцы узнают своих родителей. Наблюдения за развитием птенцов в естественных условиях показали, что птенцы иногда клюют не только клюв взрослой птицы, но и иные предметы, в том числе другие части тела родителя. Правда, большинство клевков все же нацелено на клюв, особенно в более позднем возрасте. Это позволяет предположить, что недавно вылупившиеся птенцы представляют себе родителей довольно смутно, но с возрастом, по мере накопления опыта, образ взрослой птицы становится у них все более четким. Эту проблему исследовали с помощью набора различных моделей головы и клюва (на рисунке показаны модели и относительная частота ударов по ним клювом).
Изменяя или устраняя отдельные части таких моделей, смогли выявить наиболее эффективные стимулы для пищевой реакции. Обычно модель закреплялась на стержне, который мог колебаться в такт с метрономом, так что скорость движения модели была всегда известна. В каждом опыте птенцу в случайном порядке предъявлялись пять моделей и при этом регистрировалось число клевков за определенное время (обычно за 30 секунд). Задача состояла в том, чтобы выявить наиболее эффективные стимулы для птенцов, находившихся в темноте в течение суток с момента вылупления до начала эксперимента. В первых трех опытах птенцы одинаково реагировали на все модели, кроме той, у которой не было клюва. Таким образом, показано, что новорожденные птенцы реагируют в основном на клюв родителей, а не на особенности головы (даже само наличие головы для птенцов этого возраста несущественно).
Хотя оптимальный стимул для реакции клевания, очевидно, несложен, он, видимо, должен обладать какими-то чертами, отличающими его от прочих окружающих объектов, таких как красные лапы родителей или травинки, поскольку клевки птенцов редко бывают
нацелены на посторонние предметы. Этот вопрос исследовали с помощью деревянных палочек, окрашенных в красный цвет. Палочку предъявляли птенцу в горизонтальном или вертикальном положении, причем в обоих случаях она либо была неподвижна, либо перемещалась в горизонтальном или вертикальном направлении. Любая вертикальная палочка вызывала клевание с большей частотой, чем любая горизонтальная, а из вертикальных наиболее эффективной оказалась палочка, движущаяся в горизонтальном направлении. Это хорошо согласуется с естественной ситуацией, когда вертикально
расположенный клюв взрослой птицы часто движется горизонтально перед глазами птенца.
Дальнейший анализ результатов показал, что вертикально движущийся вертикальный стимул вызывал не больше реакций, чем неподвижный вертикальный. Кроме того, и горизонтальное, и вертикальное перемещение горизонтального стимула было одинаково действенно и, как это ни странно, превосходило по эффективности неподвижный горизонтальный стимул. Наиболее осторожная интерпретация этих результатов может состоять в том, что адекватными стимулами для пищевой реакции птенцов служат два типа движения: 1) горизонтальное перемещение и 2) движение в направлении, перпендикулярном продольной оси стимула. Это позволяет объяснить, почему неподвижная вертикальная палочка оказывается не менее эффективным стимулом, чем вертикальная палочка, движущаяся вертикально. Дело в том, что в обоих случаях отсутствуют движения указанных типов. Однако в случае горизонтальной палочки вертикальное движение перпендикулярно продольной оси стимула и потому так же эффективно, как горизонтальное, и реакция здесь при обоих движениях более интенсивна, чем при полной неподвижности стимула.
На следующем этапе мы испытывали вертикальные стимулы различных диаметров, и пять скоростей движения. Палочка диаметром около 8 мм была наиболее эффективна вне зависимости от скорости движения. Скоростью, вызывающей наиболее интенсивную реакцию независимо от диаметра палочки, оказалась скорость 12 см/с. Эти результаты показывают, насколько точно реактивность в процессе эволюции подгоняется к параметрам стимулов. Толщина клюва взрослой птицы в вертикальном направлении 10,6 мм, а в горизонтальном-3,1 мм, так что средняя толщина составляет как раз около 8 мм. Кроме того, как показал анализ фильмов, снятых в естественных условиях, средняя скорость движения клюва при кормлении равна около 14,5 см/с.
Проведенный недавно эксперимент добавил еще одну особенность к образу идеального для птенцов стимула. Вертикальная палочка, находящаяся выше уровня глаз птенца, гораздо эффективнее, чем такой же стимул, предъявленный внизу. Кроме того, предпочтение отдается наклонным объектам. Такая избирательность уменьшает вероятность возникновения у птенца реакции на ноги родителей, расположенные, естественно, ниже уровня его глаз.
Теперь понятно, по крайней мере, в общих чертах, каким образом только что вылупившийся птенец отличает клюв родителей от других внешних объектов. На следующем этапе нам предстояло выяснить, изменяется ли восприятие птенца в первые несколько дней жизни. Предъявляли те же три набора по пять моделей в каждом семидневным птенцам и нашли, что их поведение заметно отличается от поведения только что вылупившихся птенцов. Более взрослые птенцы оказались чувствительными даже к незначительным изменениям формы головы и клюва. Кроме того, они четко отличали модели собственных родителей – озёрных чаек - от моделей серебристых чаек.
Итак, данные указывают на то, что недавно вылупившийся птенец лучше всего реагирует на очень простые стимулы. Хотя экспериментатор может создать модель, в качестве стимула значительно более эффективную, чем настоящая птица, в естественных условиях признаки родителей точнее соответствуют «образу» стимула для пищевой реакции, чем особенности любого другого внешнего объекта. Кроме того, по мере выкармливания птенца родителями их образ в памяти птенца детализируется. Семидневные птенцы клюют только модели достаточно сходные с взрослыми птицами того же вида..
Полагается, что вопрос о том, остаются ли все элементы стимула действительно идентичными. требует дополнительного изучения. В экспериментах Тинбергена и Пердека модель держали в руке, поэтому при ее маятникообразном движении вместе с кистью руки пятно на лбу двигалось медленнее и по более короткой дуге, чем на клюве. Кроме того, птенцу приходилось тянуться вверх, чтобы клюнуть пятно на лбу.
Можно повторить опыты Тинбергена - Пердека, но добавить еще одну модель, пятно у которой располагалось на лбу, но она закреплялась на стержне таким образом, что пятно оказывалось на таком же расстоянии от оси вращения, как и пятно на клюве у другой модели. Кроме того, экспериментальная установка была снабжена подвижным полом, высоту которого подбирали так, чтобы красное пятно на модели всегда оказывалось на уровне глаз птенца. Эту третью модель назвали «быстрой», так как пятно у нее на лбу перемещалось быстрее, чем у «медленной» модели Тинбергена и Пердека. Если гипотеза о роли скорости движения верна, то «быстрая» модель с пятном на лбу должна быть так же эффективна, как и модель с пятном на клюве.
Результаты оказались совершенно однозначными. Только что вылупившиеся птенцы одинаково охотно клевали и «быструю» модель с пятном на лбу, и обычную модель с пятном на клюве. Затем птенцы были возвращены в гнездо и подвергнуты повторным тестам на 3-й и 7-й дни жизни. Как и предполагалось, реакция на модель с пятном на клюве постепенно улучшалась по сравнению с обоими вариантами модели с пятном на лбу.
Классическая трактовка экспериментов Тинбергена - Пердека сводилась к тому, что существует некий врожденный механизм, включающийся, как полагали, при восприятии определенных довольно сложных стимулов. Эксперименты показали, что пищевое поведение птенцов чайки вначале определяется сравнительно простыми стимулами, но позже в результате научения используются более сложные стимулы. Эксперименты указывают, видимо, на то, что необходимо заново исследовать и другие случаи, которые считались примерами действия врожденных «запускающих» механизмов.
Результаты исследований подсказывают также новую интерпретацию еще одного представления, широко используемого исследователями поведения - представления о классическом условном рефлексе. В общеизвестных экспериментах И. П. Павлова до начала или во время действия безусловного стимула, вызывающего биологически адекватную реакцию, животному предъявлялся новый для него условный стимул. После некоторого числа таких сочетаний животное начинало отвечать и на отдельно предъявляемый условный стимул. В классических экспериментах Павлова звук звонка подкреплялся пищей. Через некоторое время в ответ на воздействие одного лишь звонка у животного можно было наблюдать выделение слюны.
Психологов давно интересовало, насколько полезной в обычной жизни животного может быть такая способность устанавливать связь между стимулами различных модальностей. Как в эволюции могла выработаться способность к такой форме научения, которая в естественных условиях, по-видимому, так мало используется? Результаты наводят на мысль, которую стоило бы проверить. Вспомним о том, что вначале птенец реагирует на самые простые признаки клюва взрослой птицы (безусловные стимулы), но при этом одновременно воспринимает и множество других деталей головы (условные стимулы). Под влиянием пищевого подкрепления эффективным стимулом для клевания становится все более детализированный зрительный образ.
Такого рода процесс, который можно назвать «обострением восприятия», отличается от классического условного рефлекса, наблюдаемого в лаборатории, тем, что условный и безусловный стимулы здесь физически идентичны. Не исключено, что способность к выработке классических условных рефлексов сформировалась в эволюции главным образом как механизм, лежащий в основе феномена обострения восприятия. В этом случае исследователи классических условных рефлексов, по существу, изучают побочные эффекты этого механизма. Но сейчас эти соображения - не более чем одна из возможных гипотез.
Изучали и многие другие аспекты пищевого поведения, и об одном вопросе необходимо упомянуть: речь идет о распознавании пиши. Узнают ли только что вылупившиеся птенцы пищу, когда сталкиваются с нею? Чтобы выяснить это, поставили четыре чашечки с кормом в углах небольшого ящика и пронаблюдали, как выведенные в инкубаторе птенцы первый раз в жизни находят пищу. Время, затраченное птенцом на поиски пищи, оказалось обратно пропорциональным частоте клевков. Этого и следовало ожидать, если исходить из того, что птенцы разыскивают пищу только методом проб и ошибок.
Если птенцам дать насытиться и пересадить их в другое место, пока они не проголодаются, то в повторном опыте они находят пищу гораздо быстрее. В третьем опыте необходимое время сокращается до минимума. Это нельзя объяснить увеличением частоты клевков, так как во втором в последующих опытах она лишь незначительно выше, чем в первом. Следовательно, птенцы очень быстро научаются распознавать пищу или по крайней мере ее местонахождение.
Если вылупившийся птенец вначале не умеет распознавать пищу, значит ли это, что он вынужден полагаться только на метод проб и ошибок, пока, случайно наткнувшись на корм, он не начнет быстро обучаться поискам? Наблюдения и эксперименты показывают, что есть несколько механизмов, помогающих птенцу ускорить первый поиск пиши. Запомним, что если птенец не клюет пищу, то взрослая птица обычно берет ее в клюв. При первом кормлении птенец продолжает клевать клюв родителя и после того, как срыгнутая пища падает на дно гнезда. В конце юнцов взрослая птица подбирает пищу, и птенец натыкается на нее при клевке, нацеленном на клюв родителя. Это наводит на мысль об адаптивном научении малой точности клевков только что вылупившегося птенца, так как промахи повышают вероятность того, что он при этом случайно натолкнется на пищу.
Другой механизм, помогающий птенцу быстрее научиться распознавать пищу, связан с присутствием в гнезде других птенцов. Клевки одного птенца часто бывают нацелены на белый кончик клюва другого. Обычно птенцы вылупляются с интервалом около 12 часов, и к моменту вылупления следующего птенца старший уже получает корм. Если младший пытается во время такой кормежки клевать клюв старшего, то он тоже может наткнуться на пищу. Наблюдения за гнездами подтвердили, что иногда птенцы впервые находят корм именно таким способом.
Процесс распознавания корма изучали, используя три группы птенцов серебристой чайки, выведенных в инкубаторе. Птенцы контрольной группы помещались в коробки с пищей поодиночке. Птенцы двух экспериментальных групп размещались попарно, но в одном случае второй птенец тоже никогда не видел пищи, а во втором в качестве пары подбирался птенец, ранее кормившийся в коробке. Оказалось, что птенцы, тестируемые поодиночке, дольше всех не находили пищи. Несколько меньше времени на поиск корма требовалось птенцу, компаньон которого был так же неопытен, как и он сам. Быстрее всех с задачей справлялся птенец, находившийся в коробке вместе с птенцом, уже умеющим распознавать пищу.
Такие различия нельзя объяснить только более активным «исследовательским» клеванием по принципу проб и ошибок в присутствии компаньона, так как клюв последнего, наоборот, отвлекал тестируемого птенца от таких проб. Присутствие неопытного птенца побуждало другого больше передвигаться в ящике, и это приводило к более быстрому, чем у одиночных птенцов, обнаружению пищи. Влияние обученного птенца этим не ограничивалось, так как все неопытные птенцы в его присутствии впервые находили пищу, целясь в клюв кормящегося компаньона.
Птенец чайки вступает в жизнь, обладая недостаточно координированной, неточной реакцией клевания, определяемой такими простыми признаками стимула, как форма и характер движения (мотивацией служит голод, источниками стимула - родители и другие птенцы в гнезде). Птенец вначале не способен распознавать пищу, но, промахиваясь при клевках, нацеленных на клюв другой особи, он натыкается на корм и быстро научается узнавать его. Благодаря пищевому подкреплению птенец запоминает облик родителей. В результате практики повышается точность клевков и совершенствуется оценка расстояния до цели. Птенец научается также поворачивать голову при выпрашивании пищи, что ведет к появлению отличия этой реакции от клевков, направленных непосредственно на пищу.
Обнаруженная картина позволяет думать, что и развитие других инстинктов включает элемент научения. Для формирования стереотипной видоспецифической формы поведения важно только, чтобы процесс научения у всех особей данного вида был очень сходным. На описанном примере с чайками ясно видно, что поведение нельзя осмысленно расчленить на врожденные и приобретенные формы, так же как невозможно выделить какую-либо часть поведенческого репертуара, связанную только с научением. Развитие поведения - это мозаика, обусловленная непрерывным взаимодействием организма и среды.
Источники: литература
«Жизнь животных» том 5, М. «Просвещение» - 1969
«Птицы» М. «Мир», 1983
К. Вили, В.Детье «Биология» М. «Мир», 1974
Р. Хайнд «Поведение животных» М. «Мир», 1975
«Биологический Энциклопедический Словарь», М. «Советская энциклопедия» - 1989
«Большой Энциклопедический Словарь», М. «Большая Российская Энциклопедия» - 2002