Биогеоценоз или экосистемы
Биогеоценоз
Все сообщества растений, животных, микроорганизмов, грибов находятся в теснейшей связи друг с другом, создавая неразрывную систему взаимодействующих организмов и их популяций— биоценоз, который также называют сообществом. Можно выделить сообщества любого размера и уровня. Например, в сообществе степей — сообщество луговых степей, а в нем — сообщества растений, позвоночных и беспозвоночных животных, микроорганизмов.
Среда и сообщества обмениваются веществами и энергией: из среды живые организмы поглощают вещества и энергию и возвращают их обратно в окружающую среду. Благодаря этим обменным процессам сообщество (биоценоз) и окружающая его среда представляют собой неразрывное единство, одну сложную систему. Такую систему называют экосистемой или биогеоценозом (рис. 1). В последнее время термин «экосистема» употребляется чаще.

Рис. 1. Биогеоценоз хвойного (слева) и смешанного лесов
Функциональные группы организмов в сообществе. Любое сообщество состоит из совокупности организмов, которые по типу питания можно разделить на три функциональные группы.
Зеленые растения — автотрофы. Они способны аккумулировать солнечную энергию в процессе фотосинтеза и синтезировать органические вещества. Автотрофы — это продуценты, т. е. производители органического вещества, первая функциональная группа организмов биоценоза.
Любое сообщество включает в себя также гетеротрофные организмы, которым для питания необходимы уже готовые органические вещества. Различают две группы гетеротрофов: консументы, или потребители, и редуценты, т. е. разрушители. К консументам относятся животные. Травоядные животные употребляют растительную пищу, а плотоядные — животную. К редуцентам относятся микроорганизмы — бактерии и грибы. Редуценты разлагают выделения животных, остатки мертвых растений, животных и микроорганизмов и другие органические вещества. Разрушители питаются органическими соединениями, образующимися при разложении. В процессе питания редуценты минерализуют органические отходы до воды, двуокиси углерода и минеральных элементов. Продукты минерализации вновь используются продуцентами.
Следовательно, в экосистеме пищевые и энергетические связи идут в направлении: продуценты -»- консументы -+- редуценты. Все три перечисленные группы организмов существуют в любом сообществе. В каждую группу входит множество популяций, населяющих экосистему. Только совместная работа всех трех групп обеспечивает функционирование экосистемы.
Примеры экосистем. Разные экосистемы отличаются друг от друга как по видовому составу организмов, так и по свойствам среды их обитания. Рассмотрим в качестве примеров листопадный лес и пруд.
В состав листопадных лесов входят буки, дубы, грабы, липы, клены, березы, осины, рябины и другие деревья, чья листва осенью опадает. В лесу выделяется несколько ярусов растений: высокий и низкий древесный, кустарников, трав и мохового напочвенного покрова. Растения верхних ярусов более светолюбивы и лучше приспособлены к колебаниям температуры и влажности, чем растения нижних ярусов. Кустарники, травы и мхи в лесу теневыносливы, летом они существуют в полумраке, который образуется после полного развертывания листвы деревьев. На поверхности почвы лежит подстилка, состоящая из полуразложившихся остатков, опавшей листвы, веточек деревьев и кустарников, мертвых трав (рис. 2).
Фауна листопадных лесов богата. Много норных грызунов, землероющих насекомоядных, хищников (лисица, барсук, медведь). Встречаются млекопитающие, живущие на деревьях (рысь, белка, бурундук). В группу крупных травоядных входят олени, лоси, косули. Широко распространены кабаны. Птицы гнездятся в различных ярусах леса: на земле, в кустарниках, на стволах или в дуплах и на вершинах деревьев. Много насекомых, которые питаются листьями (например, гусеницы) и древесиной (короеды). В подстилке и верхних горизонтах почвы кроме насекомых обитает громадное количество и других позвоночных животных (дождевые черви, клещи, личинки насекомых), масса грибов и бактерий.
Пример экосистемы, где средой жизни организмов служит вода,— известные всем пруды. На мелководье прудов поселяются укореняющиеся или крупные плавающие растения (камыш, кувшинки, рдесты). По всей толще воды на глубину проникновения света распространены мелкие плавающие растения, в основной массе водоросли, называемые фитопланктоном. Когда водорослей много, вода становится зеленой, как говорят, «цветет». В фитопланктоне много сине-зеленых, а также диатомовых и зеленых водорослей. Личинки насекомых, головастики, ракообразные, растительноядные рыбы питаются живыми растениями или растительными остатками, хищные насекомые и рыбы поедают разнообразных мелких животных, а крупные хищные рыбы охотятся и за растительноядными и за хищными, но более мелкими рыбами. • Организмы, разлагающие органические вещества (бактерии, жгутиковые, грибы), распространены по всему пруду, но особенно их много на дне, где накапливаются остатки мертвых растений и животных. Мы видим, как непохожи и по внешнему виду, и по видовому составу популяций экосистемы леса и пруда. Среда обитания видов разная: в лесу — воздух и почва; в пруду — воздух и вода. Однако функциональные группы живых организмов однотипны. Продуценты в лесу — деревья, кустарники, травы, мхи; в пруду — плавающие растения, водоросли и сине-зеленые. В состав консументов в лесу выходят звери, птицы, насекомые и другие беспозвоночные животные (последние населяют почву и подстилку). В пруду к консументам относятся насекомые, разные земноводные, ракообразные, растительноядные и хищные рыбы. Редуценты (грибы и бактерии) представлены в лесу наземными, в пруду — водными формами.
Эти же функциональные группы организмов существуют во всех наземных (тундры, хвойные и лиственные леса, степи, луга, пустыни) и водных (океаны, моря, озера, реки, пруды) экосистемах.

Рис. 2. Экосистема листопадного леса
ПОТОК ЭНЕРГИИ И ЦЕПИ ПИТАНИЯ
Поток энергии. Для осуществления любых жизненных процессов необходима энергия. Единственным источником энергии для зеленых растений является Солнце.
Солнечная энергия, падающая на фотосинтезирующие органы растений, аккумулируется во вновь образующихся органических соединениях. Эта энергия используется продуцентами по-разному. Часть ее тратится на дыхание, т. е. на биологическое окисление, часть запасается в виде вновь возникшей биомассы. Биомасса —- это масса организмов определенной группы или сообщества в целом.
Некоторую долю созданной продуцентами биомассы съедают травоядные животные. Хищники потребляют травоядных животных и получают долю энергии. Большая часть энергии, полученная консументами с пищей, тратится на процессы, происходящие в клетках, а также выводится с продуктами жизнедеятельности в окружающую среду. Меньшая часть энергии идет на увеличение массы тела, рост и размножение.
Часть биомассы продуцентов, не съеденная животными, отмирает, и с отмершей биомассой аккумулированная в ней энергия поступает в почву в виде растительного опада.
Растительный и животный спад (трупы + экскременты)— пища редуцентов. Определенное количество энергии запасается в биомассе редуцентов, а часть рассеивается. Редуценты отмирают, и их клетки также разлагаются. Из продуктов разложения строятся органические вещества почвы. В этих соединениях запасается энергия, которая частично тратится затем на процессы разрушения минеральных соединений.
Таким образом, энергия аккумулируется на уровне продуцентов, проходит через консументы и редуценты, входит в состав органических веществ почвы и рассеивается при разрушении ее разнообразных соединений.
Разобранный пример относится к наземным экосистемам.Подобным же образом происходят процессы и в водных экосистемах. Через любую экосистему проходит поток энергии, определенная часть которой используется каждым живым существом.
Цепи питания. Перенос энергии от ее источника (растений) через ряд организмов называют пищевой цепью. Все живые организмы связаны между собой энергетическими отношениями, поскольку являются объектами питания других организмов. Травоядные животные (потребители первого порядка) поедают растения, первичные хищники (потребители второго порядка) поедают травоядных, вторичные хищники (потребители третьего порядка) поедают хищников помельче. Таким образом, создаются пищевые цепи из продуцентов и консументов, которые на разных этапах, смыкаются с сообществом редуцентов (рис. 3).
Пищевые цепи разделяют на два типа. Один тип пищевой цепи начинается с растений и идет к растительноядным животным и далее к хищникам. Это так называемая цепь выедания (пастбищная). Другой тип начинается от растительных и животных остатков, экскрементов животных и идет к мелким животным и микроорганизмам, которые ими питаются. В результате деятельности микроорганизмов образуется полуразложившаяся масса — детрит. Такую цепь называют цепью разложения (детритной).
На суше пищевые цепи первого типа состоят обычно из 3— 5 звеньев, например: растения -> овца ->- человек — трехзвенная цепь; растения -*- кузнечики -»- ящерицы -»- ястреб — четырехзвенная цепь; растения -»- кузнечики -v лягушки-»- змеи -»--»- орел — пятизвенная цепь. Через пищевые цепи биогеоценозов суши подавляющее количество прироста растительной биомассы поступает через опад в цепи разложения.
В морях распространены такие типы цепей: фитопланктон ->- рыбы -»- хищные птицы; фитопланктон ->- мелкие ракообразные-»-рыбы, питающиеся мелкими рачками и ракообразными -»- хищные рыбы ->- хищные птицы. В водных сообществах большая часть биомассы, накопленной одноклеточными водорослями, проходит через цепь выедания и значительно меньшая включается в цепь разложения (рис. 4).
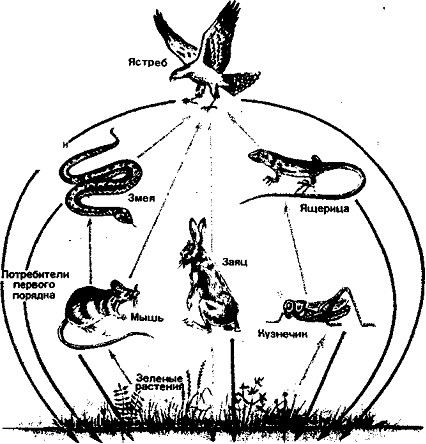
Рис. 3. Пищевые цепи в наземных экосистемах
Все типы пищевых цепей всегда существуют в сообществе таким образом, что член одной цепи является также членом другой. Соединение цепей образует пищевую сеть экосистемы. Угнетение или разрушение любого звена экосистемы с неизбежностью отразится на экосистеме в целом. Поэтому вмешиваться в жизнь экосистем надо с большой осторожностью и осмотрительностью.
Экологическая пирамида. Пищевые сети внутри каждой экосистемы имеют хорошо выраженную структуру. Она характеризуется количеством и размером организмов на каждом уровне цепи питания. При переходе с одного пищевого уровня на другой численность особей уменьшается, а их размер увеличивается. Например, в приведенной выше четырехзвенной цепи на 1 га травяной экосистемы насчитывается около 9 млн. растений (первый пищевой уровень), свыше 700 тыс. растительноядных насекомых (второй уровень), больше 350 тыс. хищных насекомых и пауков (третий уровень) и всего три птицы (четвертый уровень). Как мы видим, образуется пирамида чисел, основание которой в 3 млн. раз шире, чем вершина.
Только часть энергии, поступившей на определенный уровень биоценоза, передается организмам, находящимся на более высоком пищевом уровне. С уровня на уровень переходит около 10% энергии. Можно подсчитать, что энергия, которая доходит до пятого уровня (например, до орла в цепи: растения — кузнечики —»- лягушки - змеи -»- орел), составляет всего 0,01% энергии, поглощенной продуцентами. Таким образом, оказывается, что передача энергии с одного пищевого уровня на другой происходит с очень малым КПД. Это объясняет уменьшение числа и массы
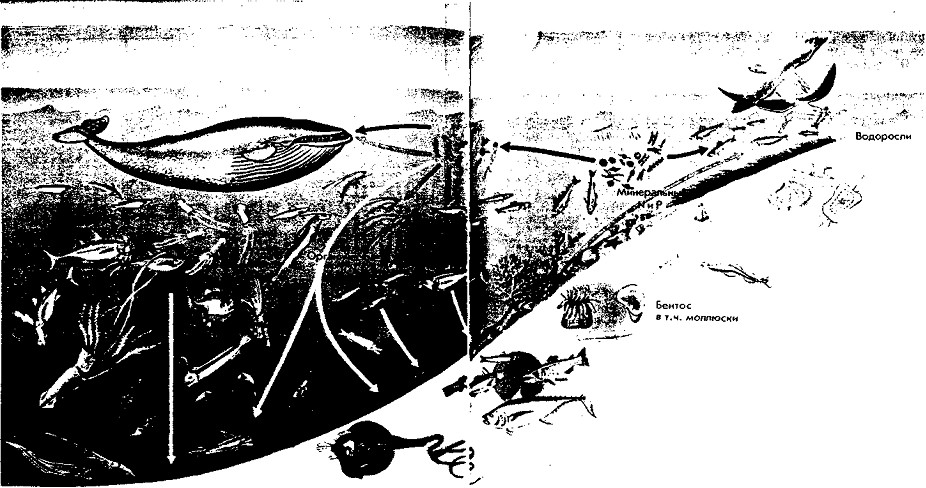
Рис. 4. Пищевые цепи в океане
Способность организмов переносить неблагоприятные условия и высокий потенциал размножения обеспечивают сохранение популяций в экосистеме, что гарантирует ее устойчивость.
Саморегуляция . Поддержание определенной численности популяций основано на взаимодействии организмов в звеньях хищник — жертва, паразит — хозяин на всех уровнях пищевых цепей. Если по каким-либо причинам один из членов пищевых цепей исчезает, то виды, питавшиеся в основном исчезнувшим видом, начинают в большем количестве поедать ту пищу, которая раньше была для них второстепенной. Вследствие подобной замены пищи численность видов-потребителей сохраняется.
Массовое размножение вида в биогеоценозе регулируется прямыми и обратными связями, существующими в пищевых цепях. Нередко благодаря хорошим погодным условиям создается высокий урожай растений, которыми питается определенная популяция травоядных животных. В связи с хорошим питанием численность популяции возрастает. Травоядные сами могут быть пищей для хищников. Чем многочисленнее жертвы, тем более обеспечен едой хищник и тем интенсивнее он размножается. Следовательно, чем больше в нынешнем году жертв, тем больше на следующий год будет хищников. Возрастание количества хищников приводит к снижению численности жертв. Снижение численности жертв ведет к тому, что размножение хищника замедляется и количество хищника и жертвы возвращается к нормальному — исходному соотношению.
Колебания количества растительной пищи, травоядных животных и хищников, питающихся этими животными, сопряжены друг с другом. Классический пример — циклы изменения численности леммингов в тундре. Раз в несколько лет на огромной территории тундры их численность резко возрастает, вслед за тем, часто за один сезон, столь же резко падает. В соответствии с этим численность песцов, лис и сов, питающихся леммингами, либо увеличивается, либо уменьшается.
Колебания численности леммингов связаны с их кормовой базой. В годы повышения численности леммингов они сильно объедают растительность. Большое количество частей растений, содержащих питательные элементы, поступает в детрит. На следующий год из-за значительного повреждения растительного покрова пищи становится меньше и питательная ценность ее уменьшается. В связи с этим рост и выживание молодых леммингов снижается. Год становится малокормным для хищников, и они почти не размножаются.
В течение последующих лет растительные остатки, богатые питательными веществами, минерализуются; питательные элементы поглощаются растениями; количество пищи- леммингов и ее питательная ценность возрастают; численность леммингов вновь стремительно идет вверх; хищники, хорошо кормясь, начинают быстро размножаться. Таким образом, в биогеоценозе популяции организмов взаимно ограничивают свою численность, благодаря чему данная экосистема существует длительное время.
Каково значение саморегуляции численности, мы понимаем особенно хорошо, сталкиваясь с явлениями, когда саморегуляция нарушается. Это обычно происходит в тех случаях, когда человек нарушает сложившуюся структуру сообществ. Примером может служить история с кроликами в Австралии.
Когда человек из Европы стал переселятся на другие континенты, он повез с собой и домашних животных, в том числе кроликов. В 1859 г. на одной из ферм Австралии выпустили 12 пар привезенных зверьков. В биогеоценозах Австралии было слишком мало хищников, чьей пищей могли бы быть кролики. Через 40 лет численность кроликов достигла нескольких сот миллионов особей. Они расселились почти по всему континенту, разоряя луга и пастбища, выедая проростки местной сосны, и нанесли урон экономике страны.
Таким образом, численность особей в природных экосистемах саморегулируется. Нарушение естественных цепей питания под воздействием антропогенного фактора, неразумное вмешательство в экосистемы может привести к неконтролируемому росту численности особей отдельных популяций и к нарушению природных экологических сообществ.
СМЕНА ЭКОСИСТЕМ
Конкретный биогеоценоз не существует вечно. Рано или поздно он сменяется другим. Смены происходят под влиянием изменения среды самими живыми организмами, при смене климатических условий, в процессе эволюции жизни на Земле, под влиянием человека.
Саморазвитие и смена экосистемы. Как пример изменения среды под влиянием самих живых организмов рассмотрим заселение растительностью скальных пород. На первых стадиях заселения большое значение имеет выветривание горных пород: разрушение, частичное растворение и изменение химических свойств минералов. Уже на самых начальных стадиях велика роль первых поселенцев: бактерий, сине-зеленых, водорослей, накипных лишайников. Сине-зеленые, свободноживущие водоросли и водоросли в составе лишайников являются продуцентами — создателями органического вещества. Многие сине-зеленые фиксируют из воздуха азот и обогащают им среду, еще мало пригодную для жизни. Лишайники выделениями органических кислот растворяют скальную породу и способствуют накоплению элементов минерального питания. Бактерии и грибы разрушают органические вещества, созданные продуцентами.
Органические вещества минерализуются не полностью. Постепенно накапливается смесь из различных органических и минеральных соединений и растительных остатков, обогащенных азотом. Создаются условия для поселения мхов и кустистых лишайников. Процесс накопления органического вещества и азота ускоряется, формируется тонкая почвенная прослойка.
Образуется примитивное сообщество, способное существовать в неблагоприятной обстановке.
Первые поселенцы хорошо приспособлены к суровым условиям голых скал — они выдерживают и сушь, и жару, и мороз. Медленно, но неуклонно они изменяют среду своего обитания и тем самым создают условия для внедрения других популяций. С приходом травянистых растений (осоки, злаки, клевер, колокольчик и др.) конкуренция за воду, свет, питательные элементы ужесточается. Пионеры-поселенцы в этой борьбе вытесняются новыми пришельцами. За травами поселяются кустарники, которые скрепляют корнями образующуюся почву. Травяно-кустарниковые сообщества сменяются лесными.
В ходе длительного развития и смены экосистемы число видов живых организмов, входящих в нее, постепенно растет. Сообщество становится более сложным, его пищевая сеть все более разветвленной. Разнообразие связей между организмами увеличивается, сообщество все полнее использует ресурсы среды. Наступает этап зрелого сообщества, наиболее хорошо приспособленного к окружающим условиям и обладающего саморегуляцией. Популяции видов в зрелом сообществе хорошо воспроизводятся и не замещаются другими видами.
Описанная смена экосистем длится тысячи лет. Однако есть смены, протекающие на глазах одного поколения людей, например зарастание мелких водоемов.
Смена экосистем под влиянием человека (антропогенного фактора).
Мощным фактором изменения экосистем является хозяйственная деятельность человека. Воздействие человека на природные экосистемы началось давно. Оно все время усиливалось вместе с увеличением населения Земли. В последнем столетии в связи с быстрым развитием промышленности, сельского хозяйства, ростом городов влияние человека приобрело решающее значение. Большие изменения происходят, например, в «зеленых зонах» . вокруг городов, которые используются для отдыха горожан. Растительность такой территории постоянно вытаптывается людьми, гуляющими по лесу, собирающими ягоды и грибы. Надземные органы растений травмируются, почва уплотняется, снижается ее способность к удержанию влаги. Все эти факторы отрицательно влияют на лесные травы, у которых корневища располагаются прямо под лесной подстилкой.
Сильное вытаптывание повреждает подрост деревьев. У кустарников и взрослых деревьев начинают сохнуть верхушки, они легко поражаются грибными заболеваниями и вредителями. В результате лес изреживается, осветляется (рис. 5). Создаются благоприятные условия для внедрения луговых трав, которые светолюбивы и меньше боятся вытаптывания, так как образуют дернину. Лесные травы становятся неконкурентоспособными и выпадают из травостоя.
Очень сильно изменяет луговые, степные и пустынные экосистемы интенсивный выпас скота. Животные поедают определенные виды трав, что приводит к распространению «не поедаемых» растений. На пастбищах разрастаются чемерица, щавель, синеголовник, полынь. Снижается обилие ценных в кормовом отношении злаков. Многие растения не успевают зацвести и дать семена. Уменьшается количество видов, упрощается сообщество. Многолетние травы замещаются однолетними растениями, корневые системы которых развиты слабее. Почва, не сдерживаемая корнями, начинает размываться потоками воды или развеваться ветром. Разрушение почвы приводит к обеднению среды питательными элементами и водой, что резко ухудшает условия жизни растений и снижает их продуктивность. Богатые разнотравные высокопродуктивные луга и степи при неумеренном выпасе превращаются в бедные пустоши.
Смена биогеоценозов под воздействием антропогенного фактора самая быстрая. Она происходит за несколько лет, а часто скачком. .К таким скачкообразным сменам относятся вырубка лесов, распашка земель с созданием агроценозов, строительство водохранилищ, когда сухопутные экосистемы превращаются в водные.
Смена экосистем под влиянием абиотических факторов. Климат земного шара неоднократно менялся. При потеплении в экосистемах вследствие естественного - отбора начинали преобладать более теплолюбивые виды растений, животных и микроорганизмов, при похолодании — холодоустойчивые. Периоды с малым количеством осадков характеризовались увеличением численности организмов, устойчивых к недостатку влаги. Периоды с обильными атмосферными осадками приводили к расцвету организмов с повышенными требованиями к содержанию влаги.
При климатогенных сменах экосистем в результате естественного отбора численность одних видов организмов сокращается, сокращается их ареал, они испытывают биологический регресс. Другие виды, оказавшиеся более устойчивыми в борьбе за существование, увеличивают численность, расширяют ареал обитания, т. е. испытывают биологический прогресс.
АГРОЦЕНОЗЫ
Структура агроценоза. Леса, тундры, степи, пустыни, реки, моря и т. д.— естественные экосистемы. Поля, огороды, сады, парки, лесные насаждения, пастбища — созданные человеком экосистемы. Их называют агроценозами.
Агроценозы — такие экосистемы, структуру и функцию которых создает, поддерживает и контролирует человек в своих интересах.
Пример агроценоза — поле пшеницы. Его растительный покров состоит в основном из растений пшеницы с примесью сорняков. Животных значительно меньше, чем в естественных экосистемах, но они есть (личинки мух, жуки, дождевые черви и др.). Иногда резко повышается численность насекомых-вредителей. Живут в норках полевки, за ними охотятся немногочисленные лисы, прилетают зерноядные и хищные птицы. Осенью урожай зерна с поля вывозят. На поле остаются солома и корневые остатки, которые разлагаются грибами и бактериями, населяющими почву.
В агроценозе, как и в любой природной экосистеме, существуют те же самые группы организмов: продуценты, консументы и редуценты. В агроценозе пшеничного поля продуцентами являются растения пшеницы и сорняков. Насекомые, птицы, полевки, лисы поедают растения или животных, т. е. принадлежат к консументам. Грибы и бактерии минерализуют органические вещества, выполняя работу редуцентов. В агроценозе складываются пищевые цепи, как и в природной экосистеме. Обязательным звеном этой пищевой цепи является человек, возделывающий поля и собирающий урожай зерна.
Энергия и питательные вещества, аккумулированные растениями, проходят по всей пищевой цепи агроценоза. Часть энергии растрачивается в процессе дыхания организмов, часть ее выносится вместе с зерном из агроценоза, часть закрепляется в органическом веществе почвы. Питательные вещества частично удаляются с урожаем, частично возвращаются в почву. Как видно из этого описания, структура и функции сообщества в агроценозе и естественном биогеоценозе похожи. Агроценоз является такой же сложной экологической системой, как лес или луг.

Рис. 5. Влияние антропогенного фактора на изменение березового леса (сравните с рис. 2)
ЭКОСИСТЕМЫ ЛЕСНОЙ ЗОНЫ УМЕРЕННЫХ ШИРОТ
Зона лесов в северном полушарии охватывает громадную территорию па материках Северной Америки и Евразии. Здесь они тянутся непрерывной полосой от побережья Тихого до Атлантического океана. В южном полушарии леса занимают небольшие площади на юге Чили и Аргентины, в Новой Зеландии, горных районах Австралии и Тасмании.
Климатические условия здесь гораздо более благоприятны для существования растительности, чем в тундрах. Так, вегетационный период длится около 4 месяцев, средняя температура июля равна +10—23 °С. Количество осадков от 250 мм в центральных частях материков до 1000 мм в год на побережьях океанов, и, что особенно важно, значительная часть из них выпадает летом. Такой режим выпадения осадков очень благоприятен для древесной растительности, и поэтому она здесь широко распространена.
В пределах зоны выделяют экосистемы темнохвойных, светлохвойных, смешанных, широколиственных и мелколиственных лесов. Темнохвойные и светлохвойные леса называют тайгой.
Древесная растительность оказывает большое воздействие на окружающую среду и изменяет ее. Густые кроны деревьев задерживают значительную часть солнечных лучей, и поэтому под пологом леса всегда сумрачно, кроны деревьев снижают скорость ветра, и в лесу царит постоянное затишье, влажность воздуха всегда высока, дождевые и талые воды хорошо впитываются рыхлой лесной подстилкой, и поэтому лесная почва хорошо увлажняется. Она же надежно предохраняет почву от пересыхания. Зимой в лесу снежный покров толще, чем на рядом расположенном луговом участке, поэтому лесная почва промерзает на незначительную глубину.
Большое воздействие оказывают деревья и на кустарниковые и травянистые растения, произрастающие в лесу. Леса умеренных широт имеют ярусное строение (рис. 6). Первый ярус составляют высокие деревья, во втором ярусе расположены кустарники, третий — образуют травянистые растения. Иногда различают даже два яруса древесных растений: ярус высоких и ярус низких деревьев. Часто мхи, покрывающие поверхность почвы зеленым ковром, образуют отдельный ярус. Корни тоже располагаются по ярусам, образуя так называемую подземную ярусность. Такое многоэтажное строение помогает растениям полнее использовать ресурсы среды обитания — свет, влагу, пищу и одновременно ослабить конкуренцию за них. Действительно, если бы все растения (деревья, кустарники и травы) брали воду и питательные элементы с динаковой
элементы с динаковой
Рис. 6. Схема ярусного строения лесных сообществ на примере
лесов Урала (по П. Л. Горчаковскому и др., 1948). Ярусы: /—ель и пихта; 2 —липа; 3 — подлесок из черемухи, рябины, жимолости, бузины, волчьего лыка; 4 — папоротник, вейник и другие травянистые растения; 5 — моховой покров.
глубины, то конкурентная борьба была бы очень острой. При ярусном расположении корневых систем деревья берут воду и пищу из наиболее глубоких горизонтов, кустарники из средних, а травянистые растения — из самых верхних. Таким образом они создают минимальные помехи ДРУГ другу.
Фитоценозы лесов значительно богаче видами, чем тундровые. Основными видами, образующими первый ярус, являются ель, пихта, лиственница, сосна, дуб, клен, ясень, липа, бук. Леса, образованные этими деревьями, приурочены к различным географическим районам в зависимости от особенностей их климатических условий.
Хвойные леса Евразии (тайга) и Северной Америки занимают территории с наиболее суровыми климатическими условиям)) — они образуют северную часть лесного пояса. Европейские и западносибирские таежные леса образованы главным образом елью и пихтой с некоторой примесью сосны и частично лиственницы. Это так называемая темнохвойная тайга. К востоку от долины Енисея расположена светлохвойная тайга. Основной породой, образующей эти леса, является лиственница.
В Европе, на Дальнем Востоке и в Северной Америке с юга к зоне хвойных лесов примыкают широколиственные леса. В Европе переход от хвойных лесов к широколиственным совершается не сразу. Между ними расположена обширная полоса смешанных лесов, которая простирается
^Лесная подстилка — слой перепревших листьев, сучьев, остатков травянистых растений, устилающих поверхность почвы.
на востоке до Урала. Эти леса образованы и хвойными, и широколиственными породами деревьев, что и отражено в их названии.
Широколиственные леса занимают обширные пространства от Британских островов па западе до Урала на востоке. Восточное Урала их сменяют мелколиственные леса. Вновь широколиственные леса появляются уже на Дальнем Востоке, на побережье Тихого океана. Как известно, чем дальше от океана расположена данная территория, тем меньше она получает влаги. Поэтому в глубинных частях материков Евразии и Северной Америки годовое количество осадков значительно меньше, чем в приокеанических районах. В зависимости от этого распределяется и растительность широколиственных лесов. В Западной Европе широко распространены влаголюбивые буковые леса, которые восточнее сменяются более сухолюбивыми дубовыми. Вместе с дубом здесь произрастают граб, клен, ясень, липа, вяз, ильм.
Мелколиственные леса Сибири образованы так называемыми мелколиственными породами — березой и осиной.
Притихоокеанские широколиственные леса востока Азии значительно богаче видами, чем европейские. Они образованы видами дуба, ореха, клена, липы, ясеня, граба, вяза. В южной части территории встречаются также пальмы, саговники и др.
Побережье Атлантического океана Северной Америки занято буковыми лесами, а в более континентальных частях распространены дубовые леса. Особенностью североамериканских широколиственных лесов является разнообразие древесных пород, которые их слагают. Так, если европейские широколиственные леса образованы не более чем 10—15 видами, то только в буковых лесах Северной Америки произрастает более 40 видов деревьев.
Второй ярус, или. как еще его называют, подлесок, в широколиственных лесах Европы образуют такие виды, как орешник, рябина, жимолость, бересклет бородавчатый. В ярусе травянистых растения широко распространены мятлики, коротконожка, осоки, копытень, сныть, медуница, колокольчики и другие виды. В таежных лесах чаще встречаются такие виды, как грушанки, брусника, черника, багульник и др.
В широколиственных лесах умеренных широт встречаются лианы: хмель, дикий виноград, плющ, ломонос. Очень интересной особенностью этих лесов является наличие группы эфемероидов. Эфемероидами называют многолетние растения, развивающиеся только весной, до распускания листьев на деревьях. К тому моменту, когда па деревьях первого яруса распустятся листья и в лесу воцарится зеленый сумрак, эфемероиды уже полностью заканчивают цикл развития. В конце мая — начале июня они исчезают из состава травостоя и сохраняются в почве до следующей весны в виде луковиц, клубней, корневищ и т. д. Эфемероиды всем хорошо известны. Как только сойдет снег, на проталинах появляются первые весенние цветы — лазоревые подснежники, желтые ветреницы, чистяк, лиловые хохлатки, розовые зубянки и др., которые местами устилают землю сплошным разноцветным ковром. Но уже через две-три недели, когда на деревьях распустятся листья, они полностью отцветают и исчезают из состава травостоя.
Развитие весенних эфемероидов, как и ярусное строение, является приспособлением к более полному использованию условий местообитания: снег уже сошел, света и тепла достаточно, а вегетация основных растений еще не началась и, если бы не было эфемероидов, благоприятное время использовалось бы биоценозом не полностью.
Леса дают более разнообразные условия для обитания животных, чем тундра: они могут питаться хвоей, листьями деревьев, их почками,, семенами, корой, древесиной, спадом, корнями, пыльцой и нектаром, стеблями, побегами, листьями травянистых растений и т. д. Лес предоставляет множество удобных укрытий и мест для устройства гнезд в кронах деревьев, под корой, под упавшими стволами деревьев, в старых пнях, в почве. Поэтому и фауна лесов значительно богаче тундровой.
В лесных экосистемах основным потребителем живых частей растений являются лось, заяц-беляк, косуля, изюбр (на Дальнем Востоке), серна, зубр, рыжая полевка, в приречных лесах бобр и многие другие. Из них листьями, ягодами, сережками, цветками, хвоей питаются рябчик, глухарь, тетерев (из тетеревиных). Живыми частями растений питается многочисленная армия насекомых, например гусеницы различных бабочек. Среди них есть и очень опасные вредители, такие, как гусеницы сибирского шелкопряда и непарного шелкопряда. Первая питается хвоей, а гусеница непарного шелкопряда — листьями деревьев. Сибирский шелкопряд распространен в основном в азиатской части, а непарный шелкопряд — в европейской части континента. Эти вредители в иные годы по неясным причинам размножаются в огромных количествах и способны уничтожить кроны деревьев на обширных территориях. Нападение гусениц наносит лесам не меньший урон, чем пожары. Многочисленны также ложногусеницы пилильщиков, листоеды и их гусеницы, майские жуки и другие хрущи. Корой и тканями деревьев питаются короеды, личинки усачей, златок. Цикадовые, тли, листоблошки питаются соками живых тканей побегов и корней; личинки долгоносиков, майского жука и другие выедают корни.
Многие растения лесов умеренных широт размножаются семенами и поэтому регулярно и обильно плодоносят. Их плоды (желуди, орехи, мелкие орешки и семена) содержат большое количество жиров и других питательных веществ. Поэтому в лесах широко распространены семеноядные животные. Среди крупных млекопитающих типичных семеноедов нет. Можно упомянуть только дикого кабана, в рационе которого заметное место занимают желуди дуба. Из более мелких животных многочисленны белки, лесные и желтогорлые мыши, сони, рыжая полевка. Этим животным семена служат основным кормом. Среди птиц наиболее известные семеноеды — это тетеревиные, сойки, клестеловик. Кроме того, заметными семеноедами являются некоторые дятлы, например большой пестрый дятел. В зимнее время он питается семенами хвойных деревьев. Некоторый вред, наносимый дятлами вследствие уничтожения семян, с лихвой окупается тем, что в остальное время года они активно уничтожают таких опасных вредителей леса, как короеды, слоники, гусеницы шелкопрядов. Не случайно в народе дятла называют доктором леса.
Из насекомых многие виды долгоносиков специализировались на поедании семян, например: желудевый долгоносик, орешниковый долгоносик, шишковая смолевка и др. Так, желудевый долгоносик в иные годы уничтожает до 70—80 % урожая желудей.
Большинство деревьев в лесах умеренных широт относится к апемофильным, т. е. ветроопыляемым. Из древесных растений наших лесов только липа и клен приспособлены к опылению насекомыми, в основном пчелами. Травянистые растения, наоборот, в подавляющем большинстве опыляются насекомыми. Исключение составляют злаки, осоки. Среди насекомых много видов антофилов («любителей цветков»). Антофилы питаются пыльцой или нектаром цветов и попутно опыляют их. Наиболее широко распрострапенными антофилами являются пчелы и шмели, бабочки, различные мухи, жуки-наездники.
В экосистемах широколиственных лесов есть группа растений, потерявших способность к фотосинтезу и паразитирующих па других растениях. Это петров крест, повилика из высших цветковых растений. Паразиты на растениях и некоторые виды грибов, например трутовик, широко встречающийся в наших лесах, а также вирусы.
Не потребленная часть первичной продукции отмирает и переходит в опад, переработкой и разложением которого занята огромная армия почвенных беспозвоночных, бактерий, грибов и актиномицетов. Среди них преобладает биомасса дождевых червей. Переработкой растительного опада заняты также личинки типулид и других двукрылых, личинки щелкунов, хрущей, очень мелкие (менее одного миллиметра) нематоды из класса круглых червей, панцирные клещики и бескрылые насекомые — коллемболы. Панцирные клещики коллемболы и по численности, и по биомассе преобладают среди мелких членистоногих, питающихся растительным опадом. Поэтому их роль в экосистемах лесов очень большая.
Почвенные беспозвоночные производят первичную «грубую» переработку опада. В таком виде опад поступает в «распоряжение» бактерий, микроскопических грибов и актиномицетов, которые доводят разложение органической массы до формы, доступной растениям.
Довольно разнообразна группа хищных животных. Из крупных млекопитающих сюда относятся в первую очередь волк, жертвами которого являются лось, косуля, бобр, заяц-беляк. В некоторых районах распространена самая крупная кошка умеренных широт — рысь, а также лисица, горностай, ласка, хорек, еж, крот, землеройка. Эти животные являются «чистыми» хищникам. т. е. они питаются исключительно животной пищей. Кроле того, некоторые хищники в зависимости от времени года и наличия пищи могут переключаться с животной пищи на растительную и наоборот. Таков, например, бурый медведь. Наиболее характерными пернатыми хищниками являются совы, филины, ястреба, соколы. Основная их пища — различные птицы и грызуны. Фактически хищниками по типу питания являются и многочисленные насекомоядные певчие птицы: славки (садовая, черноголовая, серая), синицы (большая, лазоревка, московка, гаичка), дрозды (рябинник, певчий, белобровик, деряба и др.), зяблик, горихвостка, кукушка, крапивник, мухоловки (пеструшка, малая и др.). Некоторые из них могут временно переходить на другой тип питания, например, поедать ягоды при отсутствии насекомых.
Из хищных членистоногих наиболее многочисленны пауки. Многочисленны также муравьи. Однако в отличие от пауков муравьи не являются «чистыми» (облигатными ') хищниками. Они относятся к группе так называемых факультативных 2 хищников. Значительную часть их рациона составляют семена, ягоды и т. д. Интересно, что среди' растений есть виды, специально приспособившиеся к распространению насекомыми. Это чистотел, некоторые фиалки, хохлатка, растение-паразит петров крест, гусиный лук и многие другие. Эти растения объединяются в группу мирмекохоров, т. е. «распространяемых муравьями». Муравьи питаются семенами, которые растаскивают, при этом часть из них неизбежно теряется, и таким образом растение распространяется на еще не занятые участки. Заметными хищниками являются осы. Из хищников, обитающих в подстилке, многочисленны литобииды, геофилиды, некоторые нематоды. Есть хищные виды и среди простейших. Это амебы и инфузории.
Личинки грибных комариков, моллюски, некоторые жуки, питающиеся в основном грибами, хотя и не являются хищниками в точном смысле этого слова, тем не менее в экосистемах лесов относятся к уровню хищников, точнее гетеротрофов, а не растительноядных животных, так как грибы существуют за счет готовых органических веществ.
Мы кратко рассмотрели основные группы организмов, входящих в состав пищевых цепей экосистем лесов умеренных шпрот. Сравнивая их с экосистемами тундр, мы могли убедиться, что лесные экосистемы устроены намного сложнее, чем тундровые. Они включают большее число видов, причем довольно большая их часть имеет смешанный тип питания (бурый медведь, кабан, тетеревиные).
10блигатные (обязательные) хищники — животные, питающиеся исключительно пищей животного происхождения. Как правило, облигатные хищники падалью не питаются.
2 Факультативные хищники — это животные, которые по характеру питания относятся к хищникам, но в их рационе заметное место занимает также и растительная пища.